Ląstelė bendrauja su aplinka. Biologinė evoliucija
Organizmo ryšys su aplinka fizikiniu ir cheminiu požiūriu yra atvira sistema, tai yra sistema, kurioje vyksta biocheminiai procesai. Pradinės medžiagos ateina iš aplinkos, o medžiagos, kurios taip pat nuolat susidaro, išnešamos į lauką. Pusiausvyra tarp daugiakrypčių reakcijų produktų greičio ir koncentracijos organizme yra sąlyginė, įsivaizduojama, nes medžiagų priėmimas ir pašalinimas nesiliauja. Nuolatinis ryšys su aplinka leidžia gyvą organizmą laikyti atvira sistema.
Visoms gyvoms ląstelėms energijos šaltinis yra Saulė. Augalų ląstelės chlorofilo pagalba fiksuoja saulės spindulių energiją, panaudodamos jį asimiliacijos reakcijoms fotosintezės procese. Gyvūnų, grybų ir bakterijų ląstelės saulės energiją naudoja netiesiogiai, skaidant organines medžiagas, kurias sintetina žemiški augalai.
Kai kurios ląstelės maistinės medžiagos yra suskaidomos ląstelinio kvėpavimo metu, taip aprūpindamos energiją, reikalingą įvairioms ląstelių veiklos rūšims. Šis procesas vyksta organelėse, vadinamose mitochondrijomis. Mitochondrijos susideda iš dviejų membranų: išorinės, skiriančios organelę nuo citoplazmos, ir vidinės, sudarančios daugybę raukšlių. Pagrindinis kvėpavimo produktas yra ATP. Jis palieka mitochondrijas ir yra naudojamas kaip energijos šaltinis daugeliui cheminių reakcijų citoplazmoje ir ląstelės membranoje. Jei deguonis reikalingas ląsteliniam kvėpavimui, tai kvėpavimas vadinamas aerobiniu, bet jei reakcijos vyksta nesant deguonies, tai kalbame apie anaerobinį kvėpavimą.
Bet kokiam darbui, atliekamam ląstelėje, energija naudojama vienu ir vieninteliu pavidalu - energijos iš ATP fosfatinių ryšių pavidalu. ATP yra lengvai judantis junginys. ATP susidaro ant vidinės mitochondrijų membranos. ATP sintezuojamas visose ląstelėse kvėpuojant dėl angliavandenių, riebalų ir kitų organinių medžiagų oksidacijos energijos. Žaliųjų augalų ląstelėse pagrindinis ATP kiekis dėl saulės energijos sintetinamas chloroplastuose. Fotosintezės metu jie gamina daug kartų daugiau ATP nei mitochondrijos. ATP suyra nutrūkus fosforo-deguonies ryšiams ir išsiskiriant energijai. Tai įvyksta veikiant fermentui ATPazei ATP hidrolizės metu – pridedant vandens pašalinant fosforo rūgšties molekulę. Dėl to ATP paverčiama ADP, o jei atsiskiria dvi fosforo rūgšties molekulės, tada į AMP. Kiekvienos gramo rūgšties molekulės pašalinimo reakcija lydima 40 kJ išsiskyrimo. Tai labai didelė energijos išeiga, todėl ATP fosforo ir deguonies ryšiai paprastai vadinami makroergistiniais (didelės energijos).
ATP panaudojimas plastikinėse mainų reakcijose atliekamas sujungiant jas su ATP hidrolize. Įvairių medžiagų molekulės įkraunamos energija prijungdamos nuo ATP molekulės hidrolizės metu išsiskiriančią fosforo grupę, t.y. fosforilinant.
Fosfatų darinių ypatumas yra tas, kad jie negali išeiti iš ląstelės, nors jų „išsikrovusios“ formos laisvai praeina pro membraną. Dėl šios priežasties fosforilintos molekulės lieka ląstelėje tol, kol jos panaudojamos atitinkamose reakcijose.
Atvirkštinis ADP pavertimo ATP procesas vyksta į ADP pridedant fosforo rūgšties molekulę, išskiriant vandenį ir sugeriant didelį kiekį energijos.
Taigi ATP yra universalus ir tiesioginis ląstelių veiklos energijos šaltinis. Tai sukuria vieną ląstelių energijos telkinį ir leidžia perskirstyti bei perkelti ją iš vienos ląstelės srities į kitą.
Fosfatų grupės perkėlimas atlieka svarbų vaidmenį cheminėse reakcijose, tokiose kaip makromolekulių surinkimas iš monomerų. Pavyzdžiui, aminorūgštys gali būti sujungtos į peptidus tik prieš tai fosforilintos. Mechaniniai susitraukimo ar judėjimo procesai, ištirpusios medžiagos transportavimas prieš koncentracijos gradientą ir kiti procesai apima ATP sukauptos energijos suvartojimą.
Energijos apykaitos procesą galima pavaizduoti taip. Didelės molekulinės masės organinės medžiagos citoplazmoje fermentiniu būdu, hidrolizės būdu, paverčiamos paprastesnėmis, iš kurių jos susideda: baltymai - į aminorūgštis, poli- ir disacharidai - į monosacharidus (+ gliukozė), riebalai į glicerolį ir riebalų rūgštis. Nevyksta oksidaciniai procesai, išsiskiria mažai energijos, kuri nepanaudojama ir pereina į terminę formą. Dauguma ląstelių pirmiausia naudoja angliavandenius. Polisacharidai (augaluose – krakmolas, o gyvūnuose – glikogenas) hidrolizuojasi į gliukozę. Gliukozės oksidacija vyksta trimis etapais: glikolizė, oksidacinis dekarboksilinimas (Krebso ciklas – citrinos rūgšties ciklas) ir oksidacinis fosforilinimas (kvėpavimo grandinė). Citoplazmoje vyksta glikolizė, kurios metu viena gliukozės molekulė suskaidoma į dvi piruvo rūgšties molekules, išskiriant dvi ATP molekules. Trūkstant deguonies, piruvo rūgštis paverčiama arba etanoliu (fermentacija), arba pieno rūgštimi (anaerobiniu kvėpavimu).
Kai gyvūnų ląstelėse vyksta glikolizė, šešių anglies gliukozės molekulė skyla į dvi pieno rūgšties molekules. Šis procesas yra kelių etapų. Jį paeiliui atlieka 13 fermentų. Alkoholinės fermentacijos metu iš gliukozės molekulės susidaro dvi etanolio ir dvi CO2 molekulės.
Glikolizė yra fazė, būdinga anaerobiniam ir aerobiniam kvėpavimui, kitos dvi vyksta tik aerobinėmis sąlygomis. Oksidacijos be deguonies procesas, kurio metu išsiskiria ir panaudojama tik dalis metabolitų energijos, yra galutinis anaerobiniams organizmams. Esant deguoniui, piruvo rūgštis patenka į mitochondrijas, kur dėl daugybės nuoseklių reakcijų ji visiškai aerobiškai oksiduojama iki H2O ir CO2, kartu fosforilinant ADP į ATP. Šiuo atveju dvi ATP molekulės gaminamos glikolizės būdu, dvi – Krebso cikle, o 34 – kvėpavimo grandinė. Grynasis vienos gliukozės molekulės visiško oksidacijos į H2O ir CO2 išeiga yra 38 molekulės.
Taigi aerobiniuose organizmuose galutinis organinių medžiagų skaidymas vyksta oksiduojant jas atmosferos deguonimi iki paprastų neorganinių medžiagų: CO2 ir H2O. Šis procesas vyksta mitochondrijose. Tokiu atveju išsiskiria didžiausias laisvos energijos kiekis, kurio nemaža dalis rezervuojama ATP molekulėse. Nesunku pastebėti, kad aerobinė oksidacija suteikia ląstelei daugiausia laisvos energijos.
Dėl katabolizmo ląstelėje kaupiasi daug energijos turinčios ATP molekulės, CO2 ir vandens perteklius patenka į išorinę aplinką.
Ląstelėje gali būti laikomos kvėpavimui nereikalingos cukraus molekulės. Lipidų perteklius arba suskaidomas, po to jų skilimo produktai patenka į mitochondrijas kaip kvėpavimo substratas, arba nusėda kaip rezervai citoplazmoje riebalų lašelių pavidalu. Baltymai susidaro iš aminorūgščių, patenkančių į ląstelę. Baltymų sintezė vyksta organelėse, vadinamose ribosomomis. Kiekviena ribosoma susideda iš dviejų dalelių – didelių ir mažų: abi dalelės apima baltymų molekules ir RNR molekules.
Ribosomos dažnai yra prijungtos prie specialios membranos sistemos, susidedančios iš cisternų ir pūslelių – vadinamojo endoplazminio tinklo (ER); ląstelėse, kurios gamina daug baltymų, endoplazminis tinklas dažnai būna labai gerai išvystytas ir padengtas ribosomomis. Kai kurie fermentai yra veiksmingi tik tada, kai yra prijungti prie membranos. Čia yra dauguma lipidų sintezėje dalyvaujančių fermentų. Taigi, endoplazminis tinklas yra tarsi savotiškas ląstelių darbo stalas.
Be to, ER padalija citoplazmą į atskirus skyrius, t. y. atskiria įvairius cheminius procesus, vienu metu vykstančius citoplazmoje, ir taip sumažina tikimybę, kad šie procesai trukdys vienas kitam.
Tam tikros ląstelės pagaminti produktai dažnai naudojami už ląstelės ribų. Tokiais atvejais ribosomose susintetinti baltymai praeina per endoplazminio tinklo membranas ir supakuojami į aplink jas susidarančias membranines pūsleles, kurios vėliau atsiskiria nuo ER. Šios pūslelės, išsilyginančios ir susidėjusios viena ant kitos, kaip sukrauti blynai, sudaro būdingą struktūrą, vadinamą Golgi kompleksu arba Golgi aparatu. Būdami Golgi aparate baltymai patiria tam tikrų pokyčių. Atėjus laikui joms palikti ląstelę, membranos pūslelės susilieja su ląstelės membrana ir ištuštėja, išpildamos savo turinį, t.y. sekrecija vyksta egzocitozės būdu.
Golgi aparatas gamina ir lizosomas – membraninius maišelius, kuriuose yra virškinimo fermentų. Išsiaiškinti, kaip ląstelė gamina, pakuoja ir eksportuoja tam tikrus baltymus ir kaip „žino“, kokius baltymus turėtų pasilikti sau, yra viena patraukliausių šiuolaikinės citologijos šakų.
Bet kurios ląstelės membranos nuolat juda ir keičiasi. ER membranos lėtai juda visoje ląstelėje. Atskiros šių membranų sekcijos atsiskiria ir sudaro pūsleles, kurios laikinai tampa Golgi aparato dalimi, o vėliau egzocitozės proceso metu susilieja su ląstelės membrana.
Vėliau membranos medžiaga grąžinama į citoplazmą, kur vėl panaudojama.
LĄSTELĖ
EPITELINIO AUDINIO.
AUDINIŲ RŪŠYS.
LĄSTELĖS STRUKTŪRA IR SAVYBĖS.
PASKAITA Nr.2.
1. Ląstelės sandara ir pagrindinės savybės.
2. Audinių samprata. Audinių rūšys.
3. Epitelinio audinio sandara ir funkcijos.
4. Epitelio tipai.
Tikslas: pažinti ląstelių sandarą ir savybes, audinių tipus. Atstovauti epitelio klasifikacijai ir jo vietai kūne. Gebėti atskirti epitelio audinį pagal morfologines savybes nuo kitų audinių.
1. Ląstelė – elementari gyvoji sistema, visų gyvūnų ir augalų sandaros, vystymosi ir gyvybinės veiklos pagrindas. Mokslas apie ląstelę yra citologija (gr. cytos – ląstelė, logos – mokslas). Zoologas T. Schwannas pirmą kartą suformulavo ląstelių teoriją 1839 m.: ląstelė atstovauja pagrindiniam visų gyvų organizmų sandaros vienetui, gyvūnų ir augalų ląstelės yra panašios sandaros, gyvybės už ląstelės ribų nėra. Ląstelės egzistuoja kaip nepriklausomi organizmai (protozojus, bakterijos) ir kaip dalis daugialąsčių organizmų, kuriuose yra lytinės ląstelės, kurios tarnauja dauginimuisi, ir kūno ląstelės (somatinės), skirtingos struktūros ir funkcijos (nervinės, kaulinės, sekrecinės ir kt.). ). Iš neorganinių medžiagų ląstelėje yra daugiausia vandens (70-80%), iš organinių - baltymų (10-20%) Pagrindinės ląstelės dalys yra: branduolys, citoplazma, ląstelės membrana (citolemma).
BRANDUOLIŲ CITOPLAZMA CYTOLEMMA
Nukleoplazma – hialoplazma
1-2 branduoliai – organelės
Chromatinas (endoplazminis tinklas
KToldzhi kompleksas
ląstelių centras
mitochondrijos
lizosomos
specialus tikslas)
Inkliuzai.
Ląstelės branduolys yra citoplazmoje ir nuo jos atribotas branduolio
apvalkalas – nukleolema. Jis tarnauja kaip vieta, kur koncentruojasi genai,
kurių pagrindinė cheminė medžiaga yra DNR. Branduolys reguliuoja ląstelės formavimosi procesus ir visas jos gyvybines funkcijas. Nukleoplazma užtikrina įvairių branduolinių struktūrų sąveiką, branduoliai dalyvauja ląstelių baltymų ir kai kurių fermentų sintezėje, chromatine yra chromosomos su genais – paveldimumo nešiotojais.
Hialoplazma (gr. hyalos - stiklas) yra pagrindinė citoplazmos plazma,
yra tikroji ląstelės vidinė aplinka. Jis sujungia visas ląstelių ultrastruktūras (branduolys, organelės, inkliuzai) ir užtikrina jų cheminę sąveiką tarpusavyje.
Organelės (organelės) yra nuolatinės citoplazmos ultrastruktūros, kurios ląstelėje atlieka tam tikras funkcijas. Jie apima:
1) endoplazminis tinklas - išsišakojusių kanalų ir ertmių sistema, sudaryta iš dvigubų membranų, susijusių su ląstelės membrana. Ant kanalų sienelių yra mažyčiai kūnai – ribosomos, kurios yra baltymų sintezės centrai;
2) K. Golgi kompleksas, arba vidinis tinklinis aparatas, turi tinklelius ir turi įvairaus dydžio vakuoles (lot. vakuumas – tuščias), dalyvauja ląstelių išskyrimo funkcijoje ir lizosomų formavime;
3) ląstelės centras – citocentras susideda iš sferinio tankaus kūno – centrosferos, kurios viduje yra 2 tankūs kūnai – centrioliai, sujungti trumpikliu. Įsikūręs arčiau branduolio, dalyvauja ląstelių dalijimuisi, užtikrindamas vienodą chromosomų pasiskirstymą tarp dukterinių ląstelių;
4) mitochondrijos (gr. mitos – siūlas, chondros – grūdas) turi grūdelių, strypų, siūlų išvaizdą. Jie atlieka ATP sintezę.
5) lizosomos – pūslelės, užpildytos fermentais, kurie reguliuoja
medžiagų apykaitos procesus ląstelėje ir turi virškinimo (fagocitinį) aktyvumą.
6) specialios paskirties organelės: miofibrilės, neurofibrilės, tonofibrilės, blakstienos, gaureliai, žvyneliai, atliekantys specifinę ląstelės funkciją.
Citoplazminiai intarpai yra nestabilios formos dariniai
granulės, lašeliai ir vakuolės, kuriose yra baltymų, riebalų, angliavandenių, pigmento.
Ląstelės membrana, citolema arba plazmolema, dengia ląstelės paviršių ir atskiria ją nuo aplinkos. Jis yra pusiau pralaidus ir reguliuoja medžiagų patekimą į ląstelę ir iš jos.
Tarpląstelinė medžiaga yra tarp ląstelių. Kai kuriuose audiniuose jis yra skystas (pavyzdžiui, kraujyje), o kituose jis susideda iš amorfinės (bestruktūrinės) medžiagos.
Bet kuri gyva ląstelė turi šias pagrindines savybes:
1) medžiagų apykaita arba metabolizmas (pagrindinė gyvybės savybė),
2) jautrumas (dirglumas);
3) gebėjimas daugintis (savęs dauginimasis);
4) gebėjimas augti, t.y. padidinti ląstelių struktūrų ir pačios ląstelės dydį ir tūrį;
5) gebėjimas tobulėti, t.y. specifinių funkcijų įgijimas ląstelėje;
6) sekretas, t.y. įvairių medžiagų išsiskyrimas;
7) judėjimas (leukocitai, histiocitai, sperma)
8) fagocitozė (leukocitai, makrofagai ir kt.).
2. Audinys yra panašios kilmės), sandaros ir funkcijos ląstelių sistema. Į audinių sudėtį taip pat įeina audinių skystis ir ląstelių atliekų produktai. Audinių tyrimas vadinamas histologija (gr. histos – audinys, logos – mokymas, mokslas, atsižvelgiant į struktūros, funkcijos ir vystymosi ypatybes, išskiriami šie audinių tipai:
1) epitelinis arba integumentinis;
2) jungiamieji (vidinės aplinkos audiniai);
3) raumeningas;
4) nervingas.
Ypatingą vietą žmogaus organizme užima kraujas ir limfa – skystas audinys, atliekantis kvėpavimo, trofines ir apsaugines funkcijas.
Kūne visi audiniai yra glaudžiai susiję vienas su kitu morfologiškai
ir funkcionalus. Morfologinis ryšys atsiranda dėl to, kad skirtingos
Šie audiniai yra tų pačių organų dalis. Funkcinis ryšys
pasireiškia tuo, kad skirtingų audinių, kurie sudaro, veikla
valdžios institucijos, sutiko.
Ląsteliniai ir neląsteliniai audinių elementai gyvenimo procese
veikla susidėvi ir miršta (fiziologinė degeneracija)
ir yra atkuriami (fiziologinė regeneracija). Jei pažeistas
taip pat atkuriami audiniai (reparatyvinė regeneracija).
Tačiau šis procesas vyksta ne visiems audiniams vienodai. Epitelinis
naya, atsinaujina jungiamasis, lygiųjų raumenų audinys ir kraujo ląstelės
jie dirba gerai. Atkuriamas dryžuotas raumenų audinys
tik esant tam tikroms sąlygoms. Atkuriamas nerviniame audinyje
tik nervinės skaidulos. Nervinių ląstelių dalijimasis suaugusio žmogaus organizme
asmens tapatybė nenustatyta.
3. Epitelinis audinys (epitelis) – tai audinys, dengiantis odos paviršių, akies rageną, taip pat išklojantis visas kūno ertmes, vidinį tuščiavidurių virškinimo, kvėpavimo ir kvėpavimo organų paviršių. Urogenitalinės sistemos ir yra daugumos kūno liaukų dalis. Šiuo atžvilgiu išskiriamas integumentinis ir liaukinis epitelis.
Integruotas epitelis, kaip pasienio audinys, atlieka:
1) apsauginė funkcija, apsauganti po juo esančius audinius nuo įvairių išorinių poveikių: cheminių, mechaninių, infekcinių.
2) organizmo apykaitą su aplinka, atliekant dujų mainų plaučiuose, absorbcijos plonojoje žarnoje, medžiagų apykaitos produktų (metabolitų) išsiskyrimą;
3) sudaryti sąlygas vidaus organų judrumui serozinėse ertmėse: širdyje, plaučiuose, žarnyne ir kt.
Liaukų epitelis atlieka sekrecinę funkciją, t.y. formuoja ir išskiria specifinius produktus – sekretus, kurie naudojami organizme vykstančiuose procesuose.
Morfologiškai epitelio audinys skiriasi nuo kitų kūno audinių šiais būdais:
1) jis visada užima pasienio padėtį, nes yra ant išorinės ir vidinės kūno aplinkos ribos;
2) vaizduoja ląstelių sluoksnius – epitelio ląsteles, kurios skirtingų tipų epitelyje turi skirtingas formas ir struktūras;
3) tarp epitelio ląstelių ir ląstelių nėra tarpląstelinės medžiagos
sujungti vienas su kitu įvairiais kontaktais.
4) epitelio ląstelės išsidėsčiusios ant pamatinės membranos (apie 1 µm storio plokštelė, atskirianti ją nuo apatinio jungiamojo audinio. Pamatinė membrana susideda iš amorfinės medžiagos ir fibrilinių struktūrų;
5) epitelio ląstelės turi poliškumą, t.y. bazinės ir viršūninės ląstelių dalys turi skirtingą struktūrą;
6) epitelyje nėra kraujagyslių, todėl ląstelių mityba
atliekama maistinių medžiagų difuzijos būdu per bazinę membraną iš apatinių audinių;“
7) tonofibrilių buvimas - gijinės struktūros, suteikiančios epitelio ląstelėms tvirtumo.
4. Yra keletas epitelio klasifikacijų, kurios yra pagrįstos įvairiomis savybėmis: kilme, struktūra, funkcijomis. laisva viršūninė (lot. viršūnė – viršūnė) epitelio sluoksnio dalis . Ši klasifikacija atspindi epitelio struktūrą, priklausomai nuo jo funkcijos.
Viensluoksnį plokščiąjį epitelį organizme atstovauja endotelis ir mezotelis. Endotelis iškloja kraujagysles, limfagysles ir širdies kameras. Mezotelis dengia pilvaplėvės ertmės, pleuros ir perikardo serozines membranas. Vieno sluoksnio kubinis epitelis iškloja dalį inkstų kanalėlių, daugelio liaukų latakus ir mažus bronchus. Vieno sluoksnio prizminis epitelis turi skrandžio, plonųjų ir storųjų žarnų, gimdos, kiaušintakių, tulžies pūslės, daugelio kepenų latakų, kasos, dalių gleivinę.
inkstų kanalėlių. Organuose, kuriuose vyksta absorbcijos procesai, epitelio ląstelės turi absorbcinę ribą, kurią sudaro daugybė mikrovilliukų. Viensluoksnis daugiaeilis blakstienas epitelis iškloja kvėpavimo takus: nosies ertmę, nosiaryklę, gerklas, trachėją, bronchus ir kt.
Sluoksniuotas plokščiasis nekeratinizuojantis epitelis dengia akies ragenos išorę ir burnos ertmės bei stemplės gleivinę. Pereinamasis epitelis būdingas šlapimo nutekėjimo organams: inkstų dubeniui, šlapimtakiams, šlapimo pūslei, kurių sienelės, prisipildžius šlapimo, smarkiai ištempiamos.
Išorinės sekrecijos liaukos išskiria savo išskyras į vidaus organų ertmes arba ant kūno paviršiaus. Paprastai jie turi šalinimo latakus. Endokrininės liaukos neturi latakų ir išskiria sekretą (hormonus) į kraują ar limfą.
Trečiasis evoliucijos etapas yra ląstelės išvaizda.
Baltymų ir nukleorūgščių (DNR ir RNR) molekulės sudaro biologinę ląstelę, mažiausią gyvų būtybių vienetą. Biologinės ląstelės yra visų gyvų organizmų „statybiniai blokai“ ir juose yra visi materialūs vystymosi kodai.
Ilgą laiką mokslininkai ląstelės sandarą laikė itin paprasta. Sovietiniame enciklopediniame žodyne ląstelės sąvoka aiškinama taip: „Ląstelė yra elementari gyvoji sistema, visų gyvūnų ir augalų sandaros ir gyvybinės veiklos pagrindas“. Reikėtų pažymėti, kad terminas „elementarus“ jokiu būdu nereiškia „paprasčiausias“, priešingai, ląstelė yra unikalus fraktalinis Dievo kūrinys, stulbinantis savo sudėtingumu ir tuo pačiu išskirtiniu visų jos elementų nuoseklumu. .
Kai pavyko pažvelgti į vidų elektroninio mikroskopo pagalba, paaiškėjo, kad paprasčiausios ląstelės sandara tokia pat sudėtinga ir nesuprantama kaip ir pati Visata. Šiandien jau buvo nustatyta, kad „ląstelė yra ypatinga Visatos materija, ypatinga kosmoso materija“. Vienoje ląstelėje yra informacijos, kuri gali būti tik keliose dešimtyse tūkstančių Didžiosios sovietinės enciklopedijos tomų. Tie. ląstelė, be kita ko, yra didžiulis informacijos „biorezervuaras“.
Šiuolaikinės molekulinės evoliucijos teorijos autorius Manfredas Eigenas rašo: „Kad atsitiktinai susidarytų baltymo molekulė, gamta turėtų atlikti maždaug 10 130 bandymų ir tam išleisti tiek molekulių, kurių pakaktų 1027 m. Visatos Jei baltymas buvo sukurtas protingai, tai yra, kad kiekvieno judesio pagrįstumą būtų galima patikrinti tam tikru atrankos mechanizmu, tai užtruko tik apie 2000 bandymų. Priėjome prie paradoksalios išvados: „primityvaus“ konstravimo programa gyva ląstelė“ yra užkoduota kažkur elementariųjų dalelių lygyje“.
O kaip galėtų būti kitaip? Kiekviena ląstelė, turinti DNR, yra apdovanota sąmone, suvokia save ir kitas ląsteles, kontaktuoja su Visata, iš tikrųjų yra jos dalis. Ir nors ląstelių skaičius ir įvairovė žmogaus organizme yra nuostabūs (apie 70 trilijonų), jos visos yra panašios į save, kaip ir visi ląstelėse vykstantys procesai yra panašūs. Kaip teigia vokiečių mokslininkas Rolandas Glaseris, biologinių ląstelių dizainas yra „labai gerai apgalvotas“. Kas gerai sugalvojo?
Atsakymas paprastas: baltymai, nukleorūgštys, gyvos ląstelės ir visos biologinės sistemos yra protingo Kūrėjo kūrybinės veiklos produktas.
Kas įdomu: atominiame lygmenyje nėra skirtumų tarp organinio ir neorganinio pasaulio cheminės sudėties. Kitaip tariant, atominiame lygmenyje ląstelė sukuriama iš tų pačių elementų, kaip ir negyvoji gamta. Skirtumai randami molekuliniame lygmenyje. Gyvuose organizmuose kartu su neorganinėmis medžiagomis ir vandeniu taip pat yra baltymų, angliavandenių, riebalų, nukleino rūgščių, fermento ATP sintazės ir kitų mažos molekulinės masės organinių junginių.
Iki šiol ląstelė tiesiogine prasme buvo išardyta į atomus tyrimo tikslais. Tačiau niekada neįmanoma sukurti nors vienos gyvos ląstelės, nes sukurti ląstelę reiškia sukurti gyvos Visatos dalelę. Akademikas V.P. Kaznačejevas mano, kad „ląstelė yra kosmoplanetinis organizmas... Žmogaus ląstelės yra tam tikros eterio-torsioninių biokoliderių sistemos Šiuose biokolidatoriuose vyksta mums nežinomi procesai, vyksta kosminių srautų formų materializacija, jų kosmotransformacija, o dėl to. tai dalelės materializuojasi.
Vanduo.
Beveik 80% ląstelės masės sudaro vanduo. Biologijos mokslų daktaro S. Zenino teigimu, vanduo dėl savo klasterio struktūros yra informacinė matrica, skirta kontroliuoti biocheminius procesus. Be to, būtent vanduo yra pagrindinis „taikinys“, su kuriuo sąveikauja garso dažnio virpesiai. Korinio vandens tvarka yra tokia aukšta (artima kristalo tvarkai), kad ji vadinama skystuoju kristalu.
Voverės.
Baltymai vaidina didžiulį vaidmenį biologiniame gyvenime. Ląstelėje yra keli tūkstančiai šiai ląstelėms būdingų baltymų (išskyrus kamienines ląsteles). Gebėjimas sintetinti būtent savo baltymus yra paveldimas iš ląstelės į ląstelę ir išlieka visą gyvenimą. Ląstelės gyvavimo metu baltymai palaipsniui keičia savo struktūrą ir sutrinka jų funkcija. Šie panaudoti baltymai pašalinami iš ląstelės ir pakeičiami naujais, dėl kurių palaikoma gyvybinė ląstelės veikla.
Pirmiausia atkreipkime dėmesį į baltymų konstrukcinę funkciją, nes jie yra statybinė medžiaga, iš kurios susideda ląstelių membranos ir ląstelių organeliai, kraujagyslių sienelės, sausgyslės, kremzlės ir kt.
Baltymų signalizacijos funkcija yra labai įdomi. Pasirodo, baltymai gali tarnauti kaip signalinės medžiagos, perduodančios signalus tarp audinių, ląstelių ar organizmų. Signalizacijos funkciją atlieka hormoniniai baltymai. Ląstelės gali sąveikauti viena su kita per atstumą, naudodamos signalinius baltymus, perduodamus per tarpląstelinę medžiagą.
Baltymai taip pat atlieka motorinę funkciją. Visus judesius, kuriuos gali atlikti ląstelės, pavyzdžiui, raumenų susitraukimą, atlieka specialūs susitraukiantys baltymai. Baltymai atlieka ir transportavimo funkciją. Jie sugeba pritvirtinti įvairias medžiagas ir perkelti jas iš vienos ląstelės vietos į kitą. Pavyzdžiui, kraujo baltymas hemoglobinas prijungia deguonį ir perneša jį į visus kūno audinius ir organus. Be to, baltymai atlieka ir apsauginę funkciją. Kai į organizmą patenka svetimi baltymai ar ląstelės, jis gamina specialius baltymus, kurie suriša ir neutralizuoja svetimas ląsteles ir medžiagas. Ir galiausiai, baltymų energetinė funkcija yra ta, kad visiškai suskaidžius 1 g baltymų, išsiskiria 17,6 kJ energijos.
Ląstelių struktūra.
Ląstelė susideda iš trijų neatsiejamai susijusių dalių: membranos, citoplazmos ir branduolio, o branduolio struktūra ir funkcijos skirtingais ląstelės gyvenimo laikotarpiais skiriasi. Ląstelės gyvavimo laikotarpis apima du laikotarpius: dalijimąsi, dėl kurio susidaro dvi dukterinės ląstelės, ir laikotarpį tarp dalijimų, kuris vadinamas tarpfaze.
Ląstelės membrana tiesiogiai sąveikauja su išorine aplinka ir sąveikauja su kaimyninėmis ląstelėmis. Jį sudaro išorinis sluoksnis ir po juo esanti plazminė membrana. Paviršinis gyvūnų ląstelių sluoksnis vadinamas glycocalis. Jis bendrauja su ląstelėmis su išorine aplinka ir visomis ją supančiomis medžiagomis. Jo storis yra mažesnis nei 1 mikronas.
Ląstelių struktūra
Ląstelės membrana yra labai svarbi ląstelės dalis. Jis sujungia visus ląstelių komponentus ir apibrėžia išorinę ir vidinę aplinką.
Tarp ląstelių ir išorinės aplinkos nuolat vyksta medžiagų apykaita. Iš išorinės aplinkos į ląstelę patenka vanduo, įvairios druskos atskirų jonų pavidalu, neorganinės ir organinės molekulės. Medžiagų apykaitos produktai, taip pat ląstelėje susintetintos medžiagos: baltymai, angliavandeniai, hormonai, kurie gaminasi įvairių liaukų ląstelėse, per membraną iš ląstelės pašalinami į išorinę aplinką. Medžiagų transportavimas yra viena iš pagrindinių plazminės membranos funkcijų.
Citoplazma- vidinė pusiau skysta aplinka, kurioje vyksta pagrindiniai medžiagų apykaitos procesai. Naujausi tyrimai parodė, kad citoplazma nėra kažkoks tirpalas, kurio komponentai sąveikauja tarpusavyje per atsitiktinius susidūrimus. Tai galima palyginti su želė, kuri, reaguodama į išorinį poveikį, pradeda „virpėti“. Taip citoplazma suvokia ir perduoda informaciją.
Citoplazmoje yra branduolys ir įvairios organelės, kurias jis sujungia į vieną visumą, kuri užtikrina jų sąveiką ir ląstelės, kaip vientisos integruotos sistemos, veiklą. Branduolys yra centrinėje citoplazmos dalyje. Visa vidinė citoplazmos zona užpildyta endoplazminiu tinkleliu, kuris yra ląstelinė organelė: kanalėlių, pūslelių ir „cisternų“ sistema, kurią riboja membranos. Endoplazminis tinklas dalyvauja medžiagų apykaitos procesuose, užtikrina medžiagų pernešimą iš aplinkos į citoplazmą ir tarp atskirų tarpląstelinių struktūrų, tačiau pagrindinė jo funkcija yra dalyvavimas baltymų sintezėje, kuri vyksta ribosomose. - mikroskopiniai apvalūs kūnai, kurių skersmuo 15-20 nm. Sintetinti baltymai pirmiausia kaupiasi endoplazminio tinklelio kanaluose ir ertmėse, o vėliau yra transportuojami į organelius ir ląstelių vietas, kur jie suvartojami.
Be baltymų, citoplazmoje taip pat yra mitochondrijų, mažų 0,2–7 mikronų dydžio kūnelių, kurie vadinami ląstelių „elektrinėmis“. Redokso reakcijos vyksta mitochondrijose, aprūpindamos ląsteles energija. Mitochondrijų skaičius vienoje ląstelėje svyruoja nuo kelių iki kelių tūkstančių.
Šerdis– gyvybiškai svarbi ląstelės dalis, kontroliuoja baltymų sintezę ir per juos visus fiziologinius procesus ląstelėje. Nesidalančios ląstelės branduolyje išskiriamas branduolio apvalkalas, branduolio sultys, branduolys ir chromosomos. Per branduolio apvalkalą vyksta nuolatinis medžiagų apykaita tarp branduolio ir citoplazmos. Po branduolio apvalkalu yra branduolio sultys (pusiau skysta medžiaga), kurioje yra branduolys ir chromosomos. Branduolys yra tankus apvalus kūnas, kurio matmenys gali būti labai įvairūs – nuo 1 iki 10 μm ar daugiau. Jį daugiausia sudaro ribonukleoproteinai; dalyvauja formuojant ribosomas. Paprastai ląstelėje būna 1-3 branduoliai, kartais iki kelių šimtų. Branduolys turi RNR ir baltymų.
Atsiradus ląstelei Žemėje, atsirado Gyvybė!
Tęsinys...
1 skyrius
LĄSTELIŲ FIZIOLOGIJOS PAGRINDAI
I. Dudel
Plazmos membrana . Gyvūnų ląstelės yra apribotos plazmine membrana (1.1 pav.). Šiek tiek išsamiau aptarsime jo struktūrą, kuri yra labai panaši į daugelio tarpląstelinių membranų struktūrą. Pagrindinė membranos matrica susideda iš lipidai, daugiausia fosfatidilcholino. Šie lipidai susideda iš hidrofilinės galvos grupės, prie kurios yra prijungtos ilgos hidrofobinės angliavandenilių grandinės. Vandenyje tokie lipidai spontaniškai sudaro 4–5 nm storio dvisluoksnę plėvelę, kurioje hidrofilinės grupės yra nukreiptos į vandeninę terpę, o hidrofobinės angliavandenilių grandinės išsidėsčiusios dviem eilėmis, sudarydamos bevandenę lipidinę fazę. Ląstelių membranos yra tokio tipo lipidų dvisluoksniai, juose yra glikolipidų, cholesterolio ir fosfolipidų (1.2 pav.). Hidrofilinę glikolipidų dalį sudaro oligosacharidai. Glikolipidai visada yra išoriniame plazminės membranos paviršiuje, o molekulės oligosacharidinė dalis yra orientuota kaip į aplinką panardintas plaukas. Išsklaidytos tarp fosfolipidų beveik vienodais kiekiais, cholesterolio molekulės stabilizuoja membraną. Įvairių lipidų pasiskirstymas vidiniame ir išoriniame membranos sluoksniuose yra nevienodas ir net vieno sluoksnio viduje yra sričių, kuriose koncentruojasi tam tikros rūšies lipidai. Šis netolygus pasiskirstymas
Ryžiai. 1.1. Scheminis ląstelės brėžinys, kuriame pavaizduoti svarbiausi organeliai
tikriausiai turi tam tikrą, kol kas neaiškią, funkcinę reikšmę.
Pagrindiniai funkciniai elementai, įterpti į santykinai inertišką membranos lipidų matricą, yra voverės(1.2 pav.). Įvairiose membranose baltymai sudaro 25–75 % masės, tačiau kadangi baltymų molekulės yra daug didesnės nei lipidų molekulės, 50 % masės atitinka 1 baltymo molekulės ir 50 lipidų molekulių santykį. Kai kurie baltymai prasiskverbia pro membraną iš išorinio į vidinį paviršių, o kiti yra pritvirtinti viename sluoksnyje. Baltymų molekulės paprastai yra orientuotos taip, kad jų hidrofobinės grupės būtų įterptos į lipidinę membraną, o polinės hidrofilinės grupės, esančios membranos paviršiuje, būtų įterptos į vandeninę fazę. Daugelis baltymų, esančių išoriniame membranos paviršiuje, yra glikoproteinai; jų hidrofilinės sacharidų grupės susiduria su tarpląsteline aplinka.
Tarpląstelinių organelių membraninės sistemos .
Maždaug pusę ląstelės tūrio užima organelės, iš citozolio išskirtos membranomis. Bendras tarpląstelinių organelių membranų paviršiaus plotas yra bent 10 kartų didesnis nei plazminės membranos paviršius. Plačiausiai naudojama membranų sistema yra endoplazminis Tinklelis, atstovaujantis tinklui

Ryžiai. 1.2.Scheminis plazminės membranos vaizdas. Baltymai yra įterpti į fosfolipidų dvigubą sluoksnį, kai kurie iš jų prasiskverbia į dvigubą sluoksnį, o kiti yra tik pritvirtinti prie išorinio arba vidinio sluoksnio.
labai vingiuoti vamzdeliai arba į maišelį panašios pailgos konstrukcijos; dideli endoplazminio tinklo plotai nusagstyti ribosomomis; toks tinklelis vadinamas granuliuotu arba grubiu (1.1 pav.). Goldžio kompleksas taip pat susideda iš membranomis surištų lamelių, nuo kurių nutrūksta pūslelės arba pūslelės (1.1 pav.). Lizosomos ir peroksisomos– Tai mažos specializuotos pūslelės. Visose šiose įvairiose organelėse membranoje ir erdvėje, kurią ji apima, yra specifinių fermentų rinkinių; Organelių viduje kaupiasi specialūs medžiagų apykaitos produktai, naudojami įvairioms organelių funkcijoms atlikti.
ŠerdisIr mitochondrijos skiriasi tuo, kad kiekviena iš šių organelių yra apsupta dviejų membranų. Branduolys yra atsakingas už kinetinę metabolizmo kontrolę; sulankstyta vidinė mitochondrijų membrana yra oksidacinio metabolizmo vieta; čia dėl piruvato arba riebalų rūgščių oksidacijos sintetinamas didelės energijos junginys adenozino trifosfatas (ATP, arba ATP).
Citoskeletas . Organelius supanti citoplazma jokiu būdu negali būti laikoma amorfine; jį persmelkia citoskeletinis tinklas. Citoskeletas susideda iš mikrovamzdelių, aktino gijų ir tarpinių gijų (1.1 pav.). Mikrovamzdeliai išorinis skersmuo yra apie 25 nm; jie susidaro, kaip įprastas polimeras, dėl tubulino baltymų molekulių surinkimo. Aktino gijos – susitraukiančios skaidulos, esančios beveik membraniniame sluoksnyje ir visoje ląstelėje, daugiausia dalyvauja procesuose, susijusiuose su judėjimu. Tarpinės gijos susideda iš skirtingos cheminės sudėties blokų skirtingų tipų ląstelėse; jie sudaro įvairius ryšius tarp dviejų kitų aukščiau paminėtų citoskeleto elementų. Organelės ir plazminė membrana taip pat yra susietos su citoskeletu, kuris ne tik palaiko ląstelės formą ir organelių padėtį joje, bet ir lemia ląstelės formos pokyčius bei jos judrumą.
Citozolis . Maždaug pusę ląstelės tūrio užima citozolis. Kadangi jame yra maždaug 20% baltymų, tai labiau gelis, o ne vandeninis tirpalas. Mažos molekulės, įskaitant organines ir neorganines jonai, ištirpsta vandeninėje fazėje. Tarp ląstelės ir aplinkos (tarpląstelinės erdvės) vyksta jonų mainai; šie medžiagų apykaitos procesai bus aptarti kitame skyriuje. Jonų koncentracija tarpląstelinėje erdvėje yra palaikoma dideliu tikslumu pastoviame lygyje; kiekvieno jono viduląstelinė koncentracija taip pat turi specifinį lygį, skirtingą nuo esančio už ląstelės ribų (1.1 lentelė). Ekstraląstelinėje aplinkoje gausiausias katijonas yra Na+ ląstelėje jo koncentracija daugiau nei 10 kartų mažesnė. Priešingai, K + koncentracija yra didžiausia ląstelės viduje, ji yra mažesnė nei eilės tvarka. Didžiausias tarpląstelinės ir tarpląstelinės koncentracijos gradientas yra Ca 2+, laisvųjų jonų koncentracija ląstelės viduje yra mažiausiai 10 000 kartų mažesnė nei už jos ribų. Ne visi jonai yra ištirpę citozolyje, kai kurie iš jų yra adsorbuojami ant baltymų arba nusėda organelėse. Pavyzdžiui, Ca 2+ atveju surištų jonų yra daug daugiau nei laisvųjų. Dauguma citozolinių baltymų yra fermentai, kuriuose dalyvauja daugelis tarpinio metabolizmo procesų: glikolizė ir gliukoneogenezė, aminorūgščių sintezė arba naikinimas, baltymų sintezė ribosomose (1.1 pav.). Citozolyje taip pat yra lipidų lašelių ir glikogeno granulių, kurios tarnauja kaip svarbių molekulių atsargos.
1.1 lentelė.Vidinės ir tarpląstelinės jonų koncentracijos homeoterminių gyvūnų raumenų ląstelėse. A – - „didelės molekulinės masės ląstelių anijonai“
|
Intraląstelinė koncentracija |
Ekstraląstelinė koncentracija |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
C l – |
4 mmol/l |
C1 – |
120 mmol/l |
|
HCO 3 – |
8 mmol/l |
HCO 3 – |
27 mmol/l |
|
A – |
155 mmol/l |
Kiti katijonų |
5 mmol/l |
|
Ramybės potencialas –90 mV |
|||
1.2. Medžiagų mainai tarp ląstelės ir aplinkos
Trumpai aprašėme ląstelės struktūrą, kad galėtume panaudoti šį aprašymą apžvelgdami ląstelių fiziologijos pagrindus. Jokiu būdu ląstelė negali būti laikoma statine esybe, nes nuolat vyksta medžiagų mainai tarp įvairių tarpląstelinių skyrių, taip pat tarp skyrių ir aplinkos. Ląstelių struktūros yra dinaminėje pusiausvyroje, o ląstelių sąveika tarpusavyje ir su išorine aplinka yra būtina sąlyga palaikyti funkcionuojančio organizmo gyvybę. Šiame skyriuje apžvelgsime pagrindinius tokių mainų mechanizmus. Tolesniuose skyriuose šie mechanizmai bus taikomi nervinei ląstelei ir jos funkcijoms;
tačiau tais pačiais mechanizmais veikia ir visi kiti organai.
Difuzija.Paprasčiausias medžiagos judėjimo procesas yra difuzija. Tirpaluose (ar dujose) atomai ir molekulės juda laisvai, o koncentracijų skirtumai subalansuojami dėl difuzijos. Panagrinėkime du tūrius, užpildytus skysčiu arba dujomis (1.3 pav.), kuriuose medžiagos turi koncentracijas c 1 ir c 2 ir atskirtas A paviršiaus ploto ir storio sluoksniu d. Medžiagos srautas m per laiką t aprašyta Pirmasis Ficko difuzijos dėsnis:
dm/ dt= DA/ d ( C 1 –С 2)=DA/ dD C(1)
čia D yra difuzijos koeficientas, pastovus tam tikrai medžiagai, tirpikliui ir temperatūrai. Bendresne koncentracijos skirtumo forma dc atstumu dx
dm/dt= –D A dc/dx, (2)
srautas per A sekciją yra proporcingas koncentracijos gradientui dc/dx . Minuso ženklas atsiranda lygtyje, nes koncentracijos pokytis x kryptimi yra neigiamas.
Difuzija yra svarbiausias procesas, dėl kurio dauguma vandeninių tirpalų molekulių juda nedideliais atstumais. Tai taip pat taikoma jų judėjimui ląstelėje, nes difuzijos netrukdo membranos. Daugelis medžiagų gali laisvai difunduoti per lipidų membranas, ypač vanduo ir ištirpusios dujos, tokios kaip O 2 ir CO 2 . Tirpus riebaluose

Ryžiai. 1.3.Kiekybinės difuzijos schema. Dvi erdves skiria storio sluoksnisdir plotas A. C – didelė dalelių koncentracija kairėje tūrio dalyje, C: – maža dalelių koncentracija dešinėje dalys, rausvas paviršius– koncentracijos gradientas difuziniame sluoksnyje. Difuzinis srautas dm/dt – žr lygtis (1)
medžiagos taip pat gerai pasklinda per membranas; Tai taip pat taikoma gana mažo dydžio polinėms molekulėms, tokioms kaip etanolis ir karbamidas, o cukrus sunkiai praeina per lipidų sluoksnį. Tuo pačiu metu lipidų sluoksniai yra praktiškai nepralaidūs įkrautoms molekulėms, įskaitant net neorganinius jonus. Neelektrolitams difuzijos lygtis (1) paprastai transformuojama sujungiant membranos ir difuzuojančios medžiagos charakteristikas į vieną. parametrų pralaidumas (P):
dm/dt=P AD c.(3)
Fig. 1.4 lyginami lipidų membranos pralaidumas (P) įvairioms molekulėms.
Difuzija per membranos poras . Plazminė membrana (ir kitos ląstelės membranos) yra pralaidžios ne tik per lipidinį sluoksnį difunduojančioms medžiagoms, bet ir daugeliui jonų, cukrų, aminorūgščių ir nukleotidų. Šios medžiagos prasiskverbia pro membraną per susidariusias poras transportuojančius baltymus, panardintas į membraną. Tokių baltymų viduje yra vandens užpildytas kanalas, kurio skersmuo mažesnis nei 1 nm, per kurį gali difunduoti mažos molekulės. Jie juda koncentracijos gradientu, o jei jie turi krūvį, tada jų judėjimą kanalais taip pat reguliuoja membranos potencialas. Membraniniai kanalai yra gana selektyvūs

Ryžiai. 1.4.Dirbtinių lipidų dvisluoksnių pralaidumas įvairioms medžiagoms
Kalbant apie molekulių, kurios gali praeiti pro jas, tipą, yra, pavyzdžiui, kalio, natrio ir kalcio kanalai, kurių kiekvienas yra nepralaidus beveik visiems jonams, išskyrus konkretų. Toks selektyvumas yra nulemtas kanalo sienelių surišimo vietų krūvio arba struktūros, o tai palengvina konkrečios molekulės pernešimą ir neleidžia kitoms medžiagoms prasiskverbti per kanalą (1 pav.). 1,5, A) .
Už elgesio membraniniai jonų kanalai lengva stebėti, nes srovę, atsirandančią dėl jonų judėjimo, galima išmatuoti net ir vienam kanalui. Parodyta, kad kanalai spontaniškai ir dideliu dažniu keičia savo būseną iš atviros į uždarą. Kalio kanalui būdingi srovės impulsai, kurių amplitudė yra apie 2 pA (2 10 –12 A) ir trunka keletą milisekundžių (žr. 2.12 pav., p. 37) [3]. Per šį laikotarpį per jį praeina dešimtys tūkstančių jonų. Baltymų perėjimas iš vienos konformacijos į kitą tiriamas rentgeno spindulių difrakcija, Mössbauer spektroskopija ir branduolinio magnetinio rezonanso (BMR) metodu. Taigi baltymai yra labai dinamiškos, judrios struktūros, o per baltymą einantis kanalas yra ne tik standus, vandens pripildytas vamzdelis (1.5 pav., A), bet greitai judančių molekulinių grupių ir krūvių labirintas. Ši dinaminė kanalo charakteristika atsispindi kanalo energetinis profilis, parodyta pav. 1.5, B. Čia abscisių ašis reiškia kanalo ilgį nuo išorinio tirpalo, kurio jonų koncentracija C 0 ir potencialas 0, iki vidinio tirpalo, kurio koncentracija C 1 ir potencialas E. Ordinačių ašis

Ryžiai. 1.5.A. Baltymų, sudarančių kalio kanalą, įterptą į lipidų dvisluoksnį plazmos membranos sluoksnį, schema. Ant kanalo „sienelės“ pritvirtinti keturi neigiami krūviai. B. Scheminis kanalo energijos profilis, parodytas pav. A. Ordinačių ašyje rodomos kinetinės energijos, reikalingos kanalui praeiti, reikšmės; išilgai abscisių ašies – atstumas tarp vidinio ir išorinio membranos paviršių. Energijos minimumai atitinka teigiamai įkrautų jonų su fiksuotu neigiamu krūviu surišimo vietas kanalo sienelėje. Energijos maksimumai atitinka difuzijos kliūtis kanale. Manoma, kad kanalo baltymo konformacija spontaniškai svyruoja; energijos profilio parinktys rodomos ištisomis ir punktyrinėmis linijomis; šie svyravimai labai palengvina jonų surišimą įveikiant energijos barjerą (modifikuotas)
pateikti jonų energijos lygiai kanalų surišimo vietose; smailė grafike reiškia pralaidumo barjerą, kurį turi viršyti jonų energija, kad prasiskverbtų į kanalą, o „nusileidimas“ grafike reiškia santykinai stabilią būseną (susirišimą). Nepaisant energijos piko kliūties, jonas gali prasiskverbti pro kanalą, jei energijos profilis spontaniškai pasikeičia; taip jonas gali staiga atsidurti „kitoje energijos smailės pusėje“ ir toliau judėti į ląstelę. Priklausomai nuo jonų krūvio, dydžio ir hidratacijos laipsnio bei jo gebėjimo jungtis prie kanalo sienelių struktūrų, kanalo energetinis profilis skirtingiems jonams skiriasi, o tai gali paaiškinti atskirų kanalų tipų selektyvumą.
Jonų difuzijos pusiausvyra . Įvairių jonų difuzija membranos kanalais turėtų padėti pašalinti koncentracijų skirtumus tarp tarpląstelinės ir tarpląstelinės aplinkos. Tačiau, kaip matyti iš lentelės. 1.1, tokie skirtumai išlieka, todėl jų turi būti pusiausvyra tarp difuzijos ir kitų transportavimo procesų per membraną. Kituose dviejuose skyriuose aptariami būdai, kuriais pasiekiama ši pusiausvyra. Jonų atveju difuzijos pusiausvyrą veikia jų krūvis. Neįkrautų molekulių difuziją užtikrina koncentracijų skirtumas dc , o susilyginus koncentracijoms, pats transportas sustoja. Įkrautas daleles papildomai veikia elektrinis laukas. Pavyzdžiui, kai kalio jonas palieka ląstelę pagal koncentracijos gradientą, jis turi vieną teigiamą krūvį. Taigi tarpląstelinė aplinka tampa labiau neigiamai įkrauta, todėl membranoje atsiranda potencialų skirtumas. Tarpląstelinis neigiamas krūvis neleidžia naujiems kalio jonams išeiti iš ląstelės, o tie jonai, kurie palieka ląstelę, dar labiau padidins membranos krūvį. Kalio jonų srautas sustoja, kai elektrinio lauko veikimas kompensuoja difuzijos slėgį dėl koncentracijos skirtumo. Jonai ir toliau praeina pro membraną, bet vienodais kiekiais į abi puses. Todėl, esant tam tikram jonų koncentracijos skirtumui membranoje, yra pusiausvyros potencialas E jonų prie kurio sustoja jonų srautas per membraną. Pusiausvyros potencialą galima lengvai nustatyti naudojant Nernsto lygtys:
Ejonų= RT/ zF* lnC išeina/ C in(4)
kur R – dujų konstanta, T – absoliuti temperatūra, z – jonų valentingumas (neigiamas anijonams) C išeina - tarpląstelinė jonų koncentracija, C in - intraląstelinė jonų koncentracija, F Faradėjaus numeris. Jei į lygtį pakeisime konstantas, tai esant kūno temperatūrai (T = 310 K) kalio jonų pusiausvyros potencialas E K yra lygus:
Ek= –61 mB žurnalas /(5)
Jei [ K + out ]/[ K + in ] = 39, kaip nurodyta lentelėje. 1.1 tada
Ek= –61 m B log 39= –97 mV.
Iš tiesų, buvo nustatyta, kad visos ląstelės turi membranos potencialas;žinduolių raumenų ląstelėse jo lygis yra apie -90 mV. Priklausomai nuo sąlygų ir santykinės jonų koncentracijos, ląstelių membranos potencialas gali svyruoti nuo -40 iki -120 mV. Aukščiau pateiktame pavyzdyje pateiktame langelyje (1.1 lentelė) poilsio potencialas, lygus maždaug -90 mV, rodo, kad kalio jonų srautai per membranos kanalus yra maždaug pusiausvyroje. Tai nenuostabu, nes ramybės būsenoje greičiausiai kalio kanalų būsena yra atvira, t.y. membrana pralaidiausia kalio jonams. Tačiau membranos potencialą lemia kitų jonų srautai.
Lengvumas, kuriuo neįkrautos dalelės gali išsisklaidyti per membraną, kiekybiškai įvertinamas (3) lygtyje. Pralaidumas įkrautoms dalelėms apibūdinama šiek tiek sudėtingesne lygtimi:
P= m RT/ dF(6)
Kur m- jonų mobilumas membranoje, d - membranos storis, R, T ir F – žinomos termodinaminės konstantos. Taip nustatytas įvairių jonų pralaidumo vertes galima naudoti membranos potencialui apskaičiuoti Em , kai pro membraną vienu metu praeina kalio, natrio ir chloro jonai (su pralaidumu P K, P Na ir P Cl atitinkamai). Daroma prielaida, kad potencialas tolygiai krinta membranoje, todėl lauko stiprumas yra pastovus. Šiuo atveju tai taikoma Goldmano lygtis arba pastovaus lauko lygtis :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl ) (7)
Daugumai ląstelių membranų P K maždaug 30 kartų didesnis nei P Na (taip pat žr. 1.3 skyrių). Santykinė vertė P Cl labai skiriasi; daugeliui membranų P Cl mažas, palyginti su R K tačiau kitiems (pavyzdžiui, skeleto raumenyse) P Cl , žymiai didesnis nei P K.
Aktyvus transportas, natrio pompa . Ankstesniame skyriuje aprašoma pasyvi jonų difuzija ir atsirandantis membranos potencialas esant tam tikroms intraląstelinėms ir tarpląstelinėms jonų koncentracijoms. Tačiau dėl šio proceso jonų koncentracija ląstelės viduje nėra automatiškai stabilizuojama, nes membrana
potencialas yra šiek tiek elektronneigiamesnis nei EK, ir daug palyginti su E Na (apie +60 mV). Dėl difuzijos viduląstelinės jonų koncentracijos, bent kalio ir natrio, turi būti sulygintos su tarpląstelinėmis. Jonų gradiento stabilumas pasiekiamas per aktyvų transportavimą: membranos baltymai perneša jonus per membraną prieš elektrinius ir (arba) koncentracijos gradientus, tam sunaudodami metabolinę energiją. Svarbiausias aktyvaus transporto procesas – darbas Na/K – siurblys, kuris yra beveik visose ląstelėse;
siurblys pumpuoja natrio jonus iš ląstelės, tuo pat metu pumpuoja į ląstelę kalio jonus. Tai užtikrina mažą natrio jonų koncentraciją ląstelėje ir didelę kalio jonų koncentraciją (1.1 lentelė). Natrio jonų koncentracijos gradientas ant membranos atlieka specifines funkcijas, susijusias su informacijos perdavimu elektrinių impulsų pavidalu (žr. 2.2 skyrių), taip pat kitų aktyvių transportavimo mechanizmų palaikymu ir ląstelių tūrio reguliavimu (žr. toliau). Todėl nenuostabu, kad daugiau nei 1/3 ląstelės suvartojamos energijos išleidžiama Na/K siurbliui, o kai kuriose aktyviausiose ląstelėse jo veikimui išleidžiama iki 70 proc.
Na/K transportinis baltymas yra ATPazė. Vidiniame membranos paviršiuje jis skaido ATP į ADP ir fosfatą (1.6 pav.). Vienos ATP molekulės energija panaudojama trims natrio jonams iš ląstelės ir vienu metu dviem kalio jonams pernešti į ląstelę, t.y. iš viso per vieną ciklą iš ląstelės pašalinamas vienas teigiamas krūvis. Taigi Na/K siurblys yra elektrogeninis(sukuria elektros srovę per membraną), dėl kurios membranos potencialo elektronegatyvumas padidėja maždaug 10 mV. Transporto baltymas šią operaciją atlieka dideliu greičiu: nuo 150 iki 600 natrio jonų per sekundę. Transporto baltymo aminorūgščių seka yra žinoma, tačiau šio sudėtingo metabolinio transportavimo mechanizmas dar nėra aiškus. Šis procesas aprašomas naudojant natrio ar kalio jonų perdavimo baltymais energetinius profilius (1.5,5 pav.). Pagal šių profilių pokyčių pobūdį, susijusį su nuolatiniais transportinio baltymo konformacijos pokyčiais (procesas, kuriam reikia energijos), galima spręsti apie mainų stechiometriją: du kalio jonai pakeičiami trimis natrio jonais.
Na/K siurblys, toks pat kaip izoliuotas Na+ /K + priklausoma membranos ATPazė, specifiškai slopinama širdies glikozido ouabaino (strofantino). Kadangi Na/K siurblio veikimas yra daugiapakopė cheminė reakcija, ji, kaip ir visos cheminės reakcijos, labai priklauso nuo temperatūros, kuri

Ryžiai. 1.6.Na/K siurblio-ATPazės (panardintos į plazmos membranos lipidinį dvigubą sluoksnį) diagrama, kuri per vieną ciklą pašalina tris Na + jonus iš ląstelės prieš potencialo ir koncentracijos gradientus ir į ląstelę įneša du K jonus + . Šio proceso metu viena ATP molekulė suskaidoma į ADP ir fosfatą. Diagramoje ATPazė parodyta kaip dimeras, susidedantis iš didelio (funkcinio) ir mažo subvienetų; membranoje jis egzistuoja kaip tetrameras, sudarytas iš dviejų didelių ir dviejų mažų subvienetų
parodyta pav. 1.7. Čia rodomas natrio jonų srautas iš raumenų ląstelių, palyginti su laiku; tai praktiškai prilygsta natrio jonų srautui, kurį sukelia Na/K siurblio veikimas, nes pasyvus natrio jonų srautas prieš koncentraciją ir potencialų gradientus yra itin mažas. Atšaldžius vaistą maždaug 18°C, natrio jonų srautas iš ląstelės greitai sumažės 15 kartų, o iš karto po kaitinimo atstatomas į pradinį lygį. Šis natrio jonų srauto iš ląstelės sumažėjimas yra kelis kartus didesnis nei tas, kuris atitiktų difuzijos proceso ar paprastos cheminės reakcijos priklausomybę nuo temperatūros. Panašus poveikis pastebimas, kai dėl apsinuodijimo dinitrofenoliu (DNP) išsenka medžiagų apykaitos energija (1.7,5 pav.). Vadinasi, natrio jonų srautą iš ląstelės užtikrina nuo energijos priklausoma reakcija – aktyvus siurblys. Kita siurblio savybė, kartu su didele priklausomybe nuo temperatūros ir energijos, yra prisotinimo lygis (kaip ir visos kitos cheminės reakcijos); tai reiškia, kad didėjant pernešamų jonų koncentracijai, siurblio greitis negali didėti neribotai (1.8 pav.). Priešingai, pasyviai difuzuojančios medžiagos srautas didėja proporcingai koncentracijos skirtumui pagal difuzijos dėsnį (1 ir 2 lygtys).
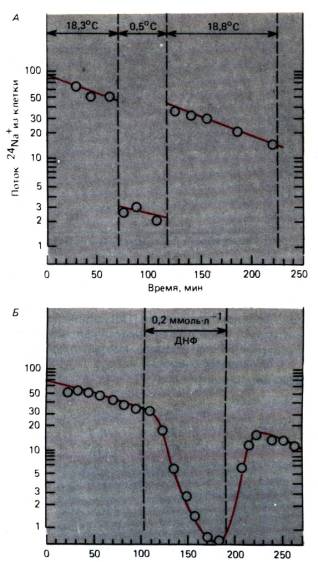
Ryžiai. 1.7. A, B. Aktyvus Na transportas + . Y ašis: radioaktyvaus 24 Na + srautas iš ląstelės (imp./min). Abscisių ašis: laikas nuo eksperimento pradžios. A. Ląstelė atšaldoma nuo 18,3 °C iki 0,5 °C; srautas Na+ iš ląstelės per šį laikotarpį yra slopinamas. B. Na + srauto iš ląstelės slopinimas dinitrofenoliu (DNP), kai koncentracija 0,2 mmol/l (modifikuota)
Be Na/K siurblio, plazminėje membranoje yra dar bent vienas siurblys – kalcio;šis siurblys pumpuoja kalcio jonus (Ca 2+) iš ląstelės ir dalyvauja palaikant itin žemą jų tarpląstelinę koncentraciją (1.1 lentelė). Kalcio siurblys labai tankiai yra raumenų ląstelių sarkoplazminiame tinkle, kuris kaupia kalcio jonus dėl ATP molekulių irimo (žr. 4 skyrių).
Na / K siurblio poveikis membranos potencialui ir ląstelių tūriui . Fig. 1.9 paveiksle pavaizduoti įvairūs membranos srovės komponentai ir parodytos jonų koncentracijos ląstelėse, kurios

Ryžiai. 1.8.Ryšys tarp molekulių transportavimo greičio ir jų koncentracijos (prie įėjimo į kanalą arba siurblio surišimo vietoje) difuzijos kanalu metu arba transportuojant pumpuojant. Pastarasis prisisotina esant didelėms koncentracijoms (maksimalus greitis, Vmax ) reikšmė x ašyje, atitinkanti pusę didžiausio siurblio greičio ( Vmax /2), yra pusiausvyros koncentracija KAM m

Ryžiai. 1.9.Diagrama, rodanti Na+ koncentracijas , K+ ir Cl – ląstelės viduje ir išorėje bei šių jonų prasiskverbimo per ląstelės membraną keliai (per specifinius jonų kanalus arba naudojant Na/K siurblį. Esant tam tikriems koncentracijos gradientams, pusiausvyros potencialai E Na, E K ir E С l – lygus nurodytam, membranos potencialas Em = – 90 mV
užtikrinti jų egzistavimą. Kalio kanalais stebima išorinė kalio jonų srovė, nes membranos potencialas yra šiek tiek elektropozityvesnis nei kalio jonų pusiausvyros potencialas. Bendras natrio kanalų laidumas yra daug mažesnis nei kalio kanalų, t.y. natrio kanalai atsidaro daug rečiau nei kalio kanalai ramybės būsenoje; tačiau į ląstelę patenka maždaug tiek pat natrio jonų, kiek kalio jonų išeina iš ląstelės, nes natrio jonams difunduoti į ląstelę reikia didelės koncentracijos ir potencialių gradientų. Na/K siurblys idealiai kompensuoja pasyvios difuzijos sroves, nes iš ląstelės išneša natrio jonus, o į ją – kalio jonus. Taigi siurblys yra elektrogeninis dėl į ląstelę ir iš jos perkeliamų krūvių skaičiaus skirtumo, kuris normaliu veikimo greičiu sukuria maždaug 10 membranos potencialą. mV yra labiau elektroneigiamas, nei tuo atveju, jei jis būtų sukurtas vien dėl pasyviųjų jonų srautų (žr. 7 lygtį). Dėl to membranos potencialas artėja prie kalio pusiausvyros potencialo, o tai sumažina kalio jonų nutekėjimą. Na veikla/K siurblį reguliuoja intraląstelinė natrio jonų koncentracija. Siurblio greitis sulėtėja, kai mažėja iš ląstelės šalinamų natrio jonų koncentracija (1.8 pav.), todėl siurblio veikimas ir natrio jonų srautas į ląstelę subalansuoja vienas kitą, išlaikant tarpląstelinę natrio koncentraciją. jonų, kurių lygis yra maždaug 10 mmol/l.
Norint išlaikyti pusiausvyrą tarp siurbimo ir pasyvių membranų srovių, reikia daug daugiau Na / K siurblio molekulių nei kalio ir natrio jonų kanalų baltymų. Kai kanalas atidarytas, per kelias milisekundes per jį praeina dešimtys tūkstančių jonų (žr. aukščiau), o kadangi kanalas dažniausiai atsidaro kelis kartus per sekundę, iš viso per šį laiką per jį praeina daugiau nei 10 5 jonų. Vieno siurblio baltymas per sekundę išjudina kelis šimtus natrio jonų, todėl plazmos membranoje turi būti apie 1000 kartų daugiau siurblio molekulių nei kanalo molekulėse. Kanalo srovių matavimai ramybės būsenoje parodė, kad 1 μm 2 membranoje yra vidutiniškai vienas kalio ir vienas natrio atviras kanalas; Iš to išplaukia, kad toje pačioje erdvėje turėtų būti apie 1000 Na/K siurblio molekulių, t.y. atstumas tarp jų yra vidutiniškai 34 nm; Siurblio baltymo, kaip kanalo baltymo, skersmuo yra 8–10 nm. Taigi membrana yra gana tankiai prisotinta pumpuojančių molekulių
Tai, kad natrio jonų srautas į ląstelę, o kalio jonai iš ląstelės, kompensuojamas veikiant siurbliui, yra dar viena pasekmė, kuri yra stabilaus osmosinio slėgio palaikymas ir pastovus tūris. Ląstelės viduje yra didelė stambių anijonų koncentracija, daugiausia baltymų (A – 1.1 lentelėje), kurie negali prasiskverbti pro membraną (arba prasiskverbia per ją labai lėtai) ir todėl yra fiksuotas komponentas ląstelės viduje. Norint subalansuoti šių anijonų krūvį, reikia vienodo katijonų skaičiaus. Dėl Na/K siurblio veikimo šie katijonai daugiausia yra kalio jonai. Reikšmingas viduląstelinių jonų koncentracijos padidėjimas galėtų įvykti tik padidėjus anijonų koncentracijai dėl C1 srauto – palei koncentracijos gradientą į ląstelę (1.1 lentelė), tačiau membranos potencialas tai atsveria. Įeinanti srovė Cl – stebimas tik tol, kol pasiekiamas chloro jonų pusiausvyros potencialas; tai pastebima, kai chloro jonų gradientas yra beveik priešingas kalio jonų gradientui, nes chloro jonai yra neigiamai įkrauti (4 lygtis). Taigi nustatoma maža intraląstelinė chloro jonų koncentracija, atitinkanti mažą ekstraląstelinę kalio jonų koncentraciją. Rezultatas yra bendro jonų skaičiaus ląstelėje apribojimas. Jei blokuojant Na/K siurblį, pavyzdžiui, esant anoksijai, membranos potencialas krenta, tai chloro jonų pusiausvyros potencialas mažėja, atitinkamai didėja chlorido jonų intraląstelinė koncentracija. Atstatant krūvių pusiausvyrą, į ląstelę patenka ir kalio jonai; padidėja bendra jonų koncentracija ląstelėje, todėl didėja osmosinis slėgis; tai priverčia vandenį į ląstelę. Ląstelė išsipučia. Šis patinimas stebimas in vivo energijos trūkumo sąlygomis.
Koncentracijos gradientas Na + kaip varomoji membranos transportavimo jėga . Na/K siurblio svarba ląstelei neapsiriboja normalių K + ir Na + gradientų stabilizavimu membranoje. Energija sukaupta membranos gradiente Na+ , dažnai naudojamas siekiant palengvinti kitų medžiagų pernešimą membranoje. Pavyzdžiui, pav. 1.10 rodo „paprastą“ Na+ ir cukraus molekules į ląstelę. Membraninis transportavimo baltymas perneša cukraus molekulę į ląstelę net esant koncentracijos gradientui, o tuo pačiu metu Na + juda išilgai koncentracijos ir potencialo gradiento, suteikdamas energiją cukrų transportavimas.Šis cukrų pernešimas visiškai priklauso nuo didelio gradiento buvimo Na+ ; jei intraląstelinė koncentracija Na+ žymiai padidėja, sustoja cukrų transportavimas. Dėl įvairių c akharov, yra įvairių simport sistemų. Amino rūgščių transportavimasį ląstelę, panašią į transportą c akharovas, parodytas fig. 1,10; tai taip pat suteikia gradientas Na+ , Yra mažiausiai penkios skirtingos simpporto sistemos, kurių kiekviena yra specializuota vienai susijusių aminorūgščių grupei.
Be paprastų sistemų, taip pat yra „antiporteriai“. Vienas iš jų, pavyzdžiui, per vieną ciklą perneša iš ląstelės vieną kalcio joną mainais į tris įeinančius natrio jonus (1.10 pav.). Energija skirta Ca 2+ transportas susidaro dėl trijų natrio jonų patekimo išilgai koncentracijos ir potencialo gradiento. Šios energijos pakanka (esant ramybės potencialui) palaikyti aukštą kalcio jonų gradientą (nuo mažiau nei 10–7 mol/L ląstelės viduje iki maždaug 2 mmol/L ląstelės išorėje).
Endo- ir egzocitozė . Tam tikroms medžiagoms, kurios patenka į ląstelę arba turi būti pašalintos

Ryžiai. 1.10.Baltymai, įterpti į lipidų dvigubą membranos sluoksnį, tarpininkauja gliukozės ir Na + simptomams į ląstelę, taip pat Ca2+/Na+ –antiportas, kuriame varomoji jėga yra Na + gradientas ant ląstelės membranos
iš jo nėra transporto kanalų; Šios medžiagos apima, pavyzdžiui, baltymus ir cholesterolį. Jie gali prasiskverbti per plazminę membraną į pūslelės, arba burbuliukai, per endo- ir egzocitozę. Fig. 1.11 paveiksle parodyti pagrindiniai šių procesų mechanizmai. Egzocitozės metu tam tikros organelės (žr. toliau) sudaro pūsleles, užpildytas medžiaga, kurią reikia pašalinti iš ląstelės, pavyzdžiui, hormonų ar tarpląstelinių fermentų. Kai tokios pūslelės pasiekia plazminę membraną, jų lipidinė membrana susilieja su ja, taip leisdama turiniui išeiti į išorinę aplinką. Esant priešingam procesui, endocitozei, plazminė membrana invaginuoja, suformuodama duobutę, kuri vėliau gilėja ir užsidaro, susiformuoja tarpląstelinė pūslelė, pripildyta tarpląstelinio skysčio ir kai kurių makromolekulių. Siekiant užtikrinti šį membranos susiliejimą ir pūslelės uždarymą, susitraukiantys citoskeleto elementai veikia kartu su pačiomis membranomis (žr. toliau). Endocitozė ne visada apima tiesiog ekstraląstelinės aplinkos patekimą į ląstelę. Ląstelės membranoje dažnai yra specifinių makromolekulių, tokių kaip insulinas ar antigenai, receptoriai, dažnai suskirstyti į specializuotas grupes. Šioms makromolekulėms prisijungus prie savo receptorių, receptorių supančioje membranos srityje įvyksta endocitozė ir makromolekulė selektyviai pernešama į ląstelę (1.12 pav., B).
Endo- ir egzocitozė ląstelėse vyksta nuolat. Membraninių medžiagų apyvartos kiekis yra reikšmingas; per 1 valandą makrofagas sugeria du kartus didesnį savo citoplazminės membranos paviršiaus plotą pūslelių pavidalu. Daugumoje ląstelių membraninės medžiagos apykaita nevyksta taip intensyviai, tačiau ji vis tiek turėtų būti reikšminga.

Ryžiai. 1.11.Egzocitozė ir endocitozė. Aukštyn: tarpląstelinė pūslelė susilieja su plazminės membranos lipidiniu dvisluoksniu sluoksniu ir atsiveria į tarpląstelinę erdvę. Šis procesas vadinamas egzocitoze. Apačioje: plazminė membrana įsiskverbia į nedidelį plotą ir išskiria tarpląstelinės medžiagos pripildytą pūslelę. Šis procesas vadinamas endocitoze
1.3. Medžiagų pernešimas ląstelėje
Endo- ir egzocitozė yra ne tik medžiagų pernešimo per ląstelės membraną procesai, bet ir membranų – pačios ląstelės struktūrinių komponentų – mainų procesai. Šiame skyriuje nagrinėjami kiti panašūs transportavimo procesai ląstelėje ir jos organelėse.

Ryžiai. 1.12. A–B. Procesų schema, įskaitant egzo- ir endocitozę. A. Granuliuotame endoplazminiame tinkle susintetintas baltymas per Golgi aparatą pernešamas į plazmos membraną, kur išskiriamas egzocitozės būdu. B. Cholesterolis, prisijungęs prie MTL (mažo tankio lipoproteinų) dalelių, prisitvirtina prie plazminės membranos, skatina endocitinės pūslelės susidarymą šioje membranos srityje ir yra transportuojamas į lizosomas, kur išsiskiria. IN. Ekstraląstelinė medžiaga, užfiksuota endocitozės metu (paveiksle Dešinėje), pernešamos per ląstelę pūslelėmis arba pūslelėmis ir išsiskiria per egzocitozę (paveiksle kairėje)
Difuzija . Natūralu, kad citozolyje koncentracijų skirtumas pašalinamas dėl difuzijos; tas pats pasakytina ir apie skysčius, esančius organelėse. Dėl didelės ištirpusių baltymų koncentracijos difuzija čia vyksta daug lėčiau nei vandenyje. Lipidų membranos – aplink ląsteles ir organelėse – yra dvimačiai skysčiai, kuriuose vyksta difuzija. Dvisluoksniame membraniniame sluoksnyje esantys lipidai difunduoja savo sluoksnyje, retai juda iš vieno į kitą. Į juos panardinti baltymai taip pat gana mobilūs; jie sukasi aplink ašį, statmeną membranai, arba difunduoja į šoną su labai skirtingomis difuzijos konstantomis, 2–10 000 kartų lėčiau nei fosfolipidai. Taigi, jei vieni baltymai lipidų sluoksnyje juda laisvai ir tokiu pat greičiu kaip ir pačios lipidų molekulės, tai kiti yra inkaruoti, t.y. gana tvirtai prijungtas prie citoskeleto. Membranoje yra „nuolatinių“ specifinių baltymų agregatų, pavyzdžiui, nervų ląstelių pre- ir postsinapsinės struktūros. Laisvai judančius baltymus galima parodyti sujungiant juos su fluorescenciniais dažais, kurių liuminescencija sukeliama trumpam apšviečiant nedidelį membranos plotą. Tokie eksperimentai rodo, kad greičiau nei per 1 minutę baltymai, prijungti prie dažų, tolygiai pasiskirsto membranoje iki 10 μm atstumu.
Aktyvus transportas organelių membranose .
Aktyvūs transportavimo procesai, kurie atlieka gyvybiškai svarbų vaidmenį plazminės membranos funkcionavime, vyksta ir ląstelės viduje, organelių membranose. Specifinis įvairių organelių turinys susidaro iš dalies dėl vidinės sintezės, iš dalies dėl aktyvaus transportavimo iš citozolio. Vienas iš pastarųjų pavyzdžių – jau minėtas Ca 2+ pompa raumenų ląstelių sarkoplazminiame tinkle. Ypač įdomu tai, kad ATP sintezės atveju mitochondrijose veikia priešingas principas tam, kas vyksta plazminės membranos ATPazės siurbliuose (1.6 pav.). ATP sintezės metu oksidacinis metabolizmas sukelia statų gradientą H+ ant vidinių membranų. Šis gradientas yra atvirkštinio aktyvaus molekulių transportavimo ciklo siurbimo proceso varomoji jėga: H + jonai juda per membraną palei gradientą, o dėl to išsiskirianti energija užtikrina ATP sintezę iš ADP ir fosfato. Gautas ATP, savo ruožtu, aprūpina ląstele energiją, įskaitant aktyvų transportavimą.
Transportavimas pūslelėse . Ląstelėje yra daug organelių ir susijusių pūslelių (1.1 pav.). Šios organelės, o ypač pūslelės, nuolat juda, pernešdamos savo turinį į kitus organelius arba į plazminę membraną. Vezikulės taip pat gali migruoti iš ląstelės membranos į organelius, kaip ir endocitoze.
Procesas baltymų sekrecija parodyta pav. 1.12, A. Baltymas sintetinamas šalia ląstelės branduolio ribosomose, susijusiose su endoplazminiu tinkleliu (vadinamasis granuliuotas arba šiurkštus endoplazminis tinklas); Patekęs į endoplazminį tinklą, baltymas supakuojamas į transportines pūsleles, kurios atsiskiria nuo organelių ir migruoja į Golgi aparatą. Čia jos susilieja su Golgi aparato cisternomis, kur baltymas modifikuojamas (t.y. paverčiamas glikoproteinu). Cisternų galuose pūslelės vėl išsiskiria. Sekretorinės pūslelės, pernešančios modifikuotą baltymą, pereina į plazmos membraną ir išskiria savo turinį egzocitozės būdu.
Kitas transportavimo kelio ląstelėje pavyzdys parodytas Fig. 1,12, B; Tai yra cholesterolio absorbcija ląstelėje. Kraujyje pernešamas cholesterolis daugiausia jungiasi su baltymais, pavyzdžiui, dalelėmis "mažo tankio lipoproteinai"(MTL). Tokios dalelės prisitvirtina prie specifinių membranų sričių, kuriose yra MTL receptorių, kur vyksta endocitozė ir MTL pernešamas į ląstelę „ribotomis“ pūslelėmis. Šios pūslelės susilieja, sudarydamos endosomas ir šio proceso metu praranda savo „apvadą“. Savo ruožtu endosomos susilieja su pirminėmis lizosomomis, kuriose vyrauja hidroliziniai fermentai, ir sudaro antrines didesnes lizosomas. Juose cholesterolis išsiskiria iš MTL dalelių ir pasklinda į citozolį, kur tampa prieinamas, pavyzdžiui, lipidų membranų sintezei. Taip pat nuo endosomų atskiriamos pūslelės, kuriose nėra MTL, kurios ypatingu būdu juda į plazminę membraną ir susilieja su ja grąžindamos membraninę medžiagą ir, tikėtina, MTL receptorius. Nuo to momento, kai MTL dalelė prisijungia prie membranos, kol cholesterolis išsiskiria iš antrinės lizosomos, praeina 10-15 minučių. MTL surišimo ir įsisavinimo, t.y. cholesterolio tiekimo į ląstelę, pažeidimai vaidina lemiamą vaidmenį vystantis rimtai ir plačiai paplitusiai ligai – aterosklerozei („arterijų sukietėjimui“).
Yra daug kitų transporto maršrutų, panašių į tuos, kurie parodyta Fig. 1.11 ir 1.12, A, kurių pagalba ląstelėje juda specifinės pūslelės. Tiksliai nežinoma, kaip jie juda, bet tikriausiai šiame procese dalyvauja citoskeleto elementai. Pūslelės gali slysti išilgai mikrovamzdelių, tokiu atveju energijos judėjimui, matyt, suteikia baltymas, susijęs su pūslelėmis, ATPazė (žr. toliau). Lieka visiškai neaišku, kiek skirtingų pūslelių, viena po kitos judančių visomis kryptimis, pasiekia savo tikslą. Akivaizdu, kad jie turi būti „pažymėti“ taip, kad būtų atpažįstami transporto sistemos ir paverčiami tikslingu judėjimu.
Transportavimas formuojant ir sunaikinant organelius . Iki šiol endo- ir egzocitozę laikėme pūslelių turinio transportavimo procesais. Yra ir dar vienas šių procesų aspektas – tai, kad tikslingas plazminės membranos pašalinimas iš vienos ląstelės paviršiaus dalies endocitozės būdu ir, priešingai, jos papildymas kitoje eksocitozės būdu, perkelia didelius membranos plotus (1 pav.). 1.12.E), suteikiant ląstelei galimybę, pavyzdžiui, suformuoti ataugą arba judėti.
Panašūs persitvarkymai būdingi ir citoskeletui, ypač mikrofilamentams ir mikrotubuliams (1.1 pav.). Mikrofilamentai pirmiausia susideda iš F-aktino baltymas, kuris gali susijungti į pluoštinius ryšulius dėl monomero polimerizacijos iš citozolio. Ryšuliai yra poliarizuoti, tai yra, jie dažnai auga tik viename gale, kaupdami naujas aktino molekules, o kitas galas yra inertiškas arba čia vyksta išardymas. Dėl šio poliarizuoto augimo mikrofilamentai efektyviai juda ir gali keistis jų tinklo struktūra. Aktino perėjimas iš depolimerizuotos būsenos (sol) į organizuotą būseną (gelis) gali įvykti labai greitai, veikiant kitiems baltymams arba pasikeitus jonų koncentracijai (žr. toliau). Taip pat yra baltymų, dėl kurių aktino gijos suyra ir susidaro trumpi fragmentai. Daugelio ląstelių plonose projekcijose – filopodijose – yra centrinis aktino pluoštas (1.1 pav.), o įvairūs filopodijų judesiai greičiausiai atsiranda dėl aktino perėjimų: polimerizacija – depolimerizacija.
Mikrovamzdeliaitaip pat dažnai patiria panašius judesius. Šių judesių mechanizmas panašus – tubulino polimerizacija iš citozolio taip, kad vienas iš mikrotubulo galų išauga, o kitas arba nesikeičia, arba ten vyksta išardymas. Taigi, mikrovamzdeliai, tinkamai pridedant arba pašalinus medžiagą, gali judėti visame citozolyje.
Aktyvūs citoskeleto judesiai . Citoskeleto struktūrų pokyčiai gali atsirasti tiek dėl aktyvių judesių, tiek dėl aukščiau aprašytų pertvarkymų. Daugeliu atvejų mikrotubulių ir aktino gijų judėjimą skatina susitraukiantys baltymai, kurie suriša siūlus ar kanalėlius ir gali juos perkelti vienas kito atžvilgiu. Voverės miozinas ir dyneinas yra visų ląstelių citozolyje palyginti didelėmis koncentracijomis; jie yra elementai, paverčiantys energiją į judėjimą specializuotose ląstelėse (raumenyse) ir organelėse (blakstienose). Raumenų ląstelėse miozinas sudaro storas gijas, orientuotas lygiagrečiai aktino gijomis. Miozino molekulė su savo „galva“ prisitvirtina prie aktino gijos ir, naudodama ATP energiją, išstumia mioziną išilgai aktino molekulės. Tada miozinas atsiskiria nuo aktino. Daugelio tokių prijungimo-atjungimo ciklų derinys lemia makroskopinį raumenų skaidulų susitraukimas(4 skyrius). Dineinas atlieka panašų vaidmenį mikrotubulių judėjime blakstienų metu (1.1 pav.). Nespecializuotų ląstelių citoplazmoje miozinas ir dyneinas nesudaro taisyklingų skaidulų, o dažniausiai nedideles molekulių grupes. Net ir tokiuose mažuose agregatuose jie gali perkelti aktino gijas arba mikrovamzdelius. Ryžiai. 1.13 paveikslas iliustruoja šį procesą, kai priešingai poliarizuotos miozino molekulės taip pat yra prijungtos prie dviejų aktino gijų, poliarizuotų skirtingomis kryptimis. Miozino galvutės lenkiasi link molekulės uodegos, sunaudodamos ATP, o dvi aktino gijos juda priešinga kryptimi, o po to miozinas nuo jų atsiskiria. Tokio pobūdžio judesiai, kurių metu ATP energija paverčiama mechaniniu darbu, gali pakeisti citoskeleto, taigi ir ląstelės, formą, taip pat užtikrinti su citoskeletu susijusių organelių transportavimą.
Aiškiausiai tarpląstelinio transportavimo procesai gali būti parodyti nervinės ląstelės aksone. Axon transportasčia išsamiai aptariamas, siekiant iliustruoti įvykius, kurie gali įvykti panašiai daugumoje ląstelių. Vos kelių mikronų skersmens aksonas gali siekti metrą ar daugiau, o baltymų judėjimas difuzijos būdu iš branduolio į distalinį aksono galą užtruktų metus. Jau seniai žinoma, kad susiaurėjus bet kuriai aksono daliai, proksimaliau esanti aksono dalis išsiplečia. Atrodo, kad išcentrinis srautas yra užblokuotas aksone. Toks srautas – greitas aksonų pernešimas gali parodyta radioaktyviųjų žymenų judėjimu, kaip parodyta Fig. 1.14. Radioaktyviai pažymėtas leucinas buvo švirkščiamas į nugaros šaknies ganglioną, o po to radioaktyvumas buvo matuojamas sėdimojo nervo 166 mm atstumu nuo neuronų ląstelių kūnų nuo 2 iki 10 valandos. Per 10 valandų radioaktyvumo pikas injekcijos vietoje šiek tiek pasikeitė. Tačiau radioaktyvumo banga išilgai aksono pasklido pastoviu maždaug 34 mm greičiu per 2 valandas arba 410 mm per dieną. Įrodyta, kad visuose homeoterminių gyvūnų neuronuose greitas aksonų pernešimas vyksta tuo pačiu greičiu, o tarp plonų, nemielinizuotų skaidulų ir storiausių aksonų, taip pat tarp motorinių ir sensorinių skaidulų pastebimų skirtumų nepastebėta. Radioaktyvaus žymens tipas taip pat neturi įtakos greito aksoninio pernešimo greičiui; žymekliai gali tarnauti kaip įvairūs radioaktyvūs

Ryžiai. 1.13.Neraumeninis miozino kompleksas, turėdamas tam tikrą orientaciją, gali prisijungti prie skirtingo poliškumo aktino gijų ir, naudodamas ATP energiją, jas išstumti vienas kito atžvilgiu.
molekulės, tokios kaip įvairios aminorūgštys, kurios yra įtrauktos į neurono ląstelės kūno baltymus. Jei analizuojame periferinę nervo dalį, kad nustatytų čia pernešamų radioaktyvumo nešėjų pobūdį, tada tokių nešiklių daugiausia randama baltymų frakcijoje, taip pat tarpininkų ir laisvųjų aminorūgščių sudėtyje. Žinodami, kad šių medžiagų savybės yra skirtingos, o jų molekulių dydžiai ypač skiriasi, pastovų transportavimo greitį galime paaiškinti tik visoms joms bendru transportavimo mechanizmu.
Aprašyta aukščiau greitas aksonų transportavimas yra anterogradinis, y., nukreiptas nuo ląstelės kūno. Buvo įrodyta, kad kai kurios medžiagos juda iš periferijos į ląstelės kūną retrogradinis transportas. Pavyzdžiui, acetilcholinesterazė šia kryptimi pernešama 2 kartus lėtesniu greičiu nei greitas aksoninis transportas. Neuroanatomijoje dažnai naudojamas žymeklis – krienų peroksidazė – taip pat juda retrogradiniu transportu. Retrogradinis transportas tikriausiai vaidina svarbų vaidmenį reguliuojant baltymų sintezę ląstelės kūne. Praėjus kelioms dienoms po aksono perpjovimo, ląstelės kūne stebima chromatolizė, o tai rodo baltymų sintezės sutrikimą. Laikas, reikalingas chromatolizei, koreliuoja su retrogradinio transportavimo iš aksono transekcijos vietos į ląstelės kūną trukme. Šis rezultatas taip pat rodo šio sutrikimo paaiškinimą – sutrinka baltymų sintezę reguliuojančios „signalinės medžiagos“ perdavimas iš periferijos. Akivaizdu, kad pagrindinės „transporto priemonės“ naudojamos greitam aksoniniam

Ryžiai. 1.14.Eksperimentas, rodantis greitą aksonų pernešimą katės sėdimojo nervo jutimo skaidulose. Tričiu pažymėtas leucinas sušvirkščiamas į nugaros šaknies ganglioną, o radioaktyvumas ganglione ir jutiminėse skaidulose matuojamas praėjus 2, 4, 6, 8 ir 10 valandų po injekcijos. (paveikslo apačioje). Autorius x ašis Nubraižytas atstumas nuo gangliono iki sėdimojo nervo sričių, kuriose atliekamas matavimas. Ordinačių ašyje tik viršutinėje ir apatinėje kreiviuose radioaktyvumas (impulsai/min.) vaizduojamas logaritmine skale. Padidėjusio radioaktyvumo „banga“. (rodyklės) juda 410 mm per dieną greičiu (ilgai)
transportas yra pūslelės (pūslelės) ir organelės, pavyzdžiui, mitochondrijos, kuriose yra medžiagų, kurias reikia transportuoti. Didžiausių pūslelių arba mitochondrijų judėjimą galima stebėti naudojant mikroskopą in vivo . Tokios dalelės daro trumpus, staigius judesius viena kryptimi, sustoja, dažnai šiek tiek pasislenka atgal arba į šoną, vėl sustoja, o tada trūkčioja pagrindine kryptimi. 410 mm/parą atitinka vidutinį anterogradinio judėjimo greitį maždaug 5 μm/s; Todėl kiekvieno atskiro judesio greitis turi būti daug didesnis, o jei atsižvelgsime į organelių, gijų ir mikrovamzdelių dydžius, tai šie judesiai iš tiesų yra labai greiti. Greitam aksoniniam transportavimui reikalinga didelė ATP koncentracija. Tokie nuodai, kaip kolchicinas, ardantis mikrovamzdelius, taip pat blokuoja greitą aksonų pernešimą. Iš to išplaukia, kad mūsų nagrinėjamame transportavimo procese pūslelės ir organelės juda išilgai mikrotubulių ir aktino gijų; šį judėjimą užtikrina maži dyneino ir miozino molekulių agregatai, veikiantys kaip parodyta Fig. 1,13, naudojant ATP energiją.
Taip pat gali būti naudojamas greitas aksonų pernešimas patologiniai procesai. Kai kurie neurotropiniai virusai (pavyzdžiui, herpeso ar poliomielito virusai) patenka į aksoną periferijoje ir retrogradiniu transportu juda į neuronų kūną, kur dauginasi ir daro toksinį poveikį. Stabligės toksinas – baltymas, kurį gamina bakterijos, kurios patenka į organizmą, kai pažeidžiama oda, sugaunamas nervų galūnėlių ir pernešamas į neuronų kūną, kur sukelia būdingus raumenų spazmus. Yra žinomi toksinio poveikio pačiam aksonų transportavimui atvejai, pavyzdžiui, pramoninio tirpiklio akrilamido poveikis. Be to, manoma, kad vitaminų trūkumo beriberi ir alkoholinės polineuropatijos patogenezė yra susijusi su greito aksonų transportavimo pažeidimu.
Be greito aksonų pernešimo ląstelėje, yra ir gana intensyvus lėtas aksonų pernešimas. Tubulinas juda palei aksoną maždaug 1 mm per dieną greičiu, o aktinas greičiau – iki 5 mm per dieną. Su šiais citoskeleto komponentais migruoja ir kiti baltymai; Pavyzdžiui, atrodo, kad fermentai yra susiję su aktinu arba tubulinu. Tubulino ir aktino judėjimo greitis apytiksliai atitinka anksčiau aprašyto mechanizmo augimo greitį, kai molekulės yra įtrauktos į aktyvųjį mikrotubulo ar mikrofilamento galą. Todėl šis mechanizmas gali būti lėto aksoninio transportavimo pagrindas. Lėto aksoninio transportavimo greitis taip pat maždaug atitinka aksonų augimo greitį, o tai, matyt, rodo apribojimus, kuriuos citoskeleto struktūra nustato antrajam procesui.
Baigiant šį skyrių, reikia pabrėžti, kad ląstelės jokiu būdu nėra statiškos struktūros, kaip matyti, pavyzdžiui, elektroninėse mikroskopinėse nuotraukose. Plazmos membrana ir ypač organelės nuolat greitai juda ir nuolat restruktūrizuojasi; tai vienintelė priežastis, kodėl jie gali veikti. Be to, tai ne paprastos kameros, kuriose vyksta cheminės reakcijos, o labai organizuoti membranų ir pluoštų konglomeratai, kurioje reakcijos vyksta optimaliai organizuota seka.
1.4. Ląstelių funkcijų reguliavimas
Atskiros ląstelės, kaip funkcinio vieneto, palaikymą pirmiausia reguliuoja branduolys; tokių reguliavimo mechanizmų tyrimas yra ląstelių biologijos ir biochemijos dalykas. Tuo pačiu metu ląstelės turi keisti savo funkcijas, atsižvelgdamos į aplinkos sąlygas ir kitų organizmo ląstelių poreikius, t.y. jos tarnauja kaip funkcinio reguliavimo objektai. Žemiau trumpai apžvelgsime, kaip šie reguliavimo poveikiai veikia plazmos membraną ir kaip jie pasiekia tarpląstelinius organelius.
Reguliuojantis poveikis ląstelių membranai
Membranos potencialas . Daugeliu atvejų ląstelių funkcijų reguliavimas atliekamas keičiant membranos potencialą. Vietiniai potencialo pokyčiai galimi, kai: 1) per membraną teka srovė iš gretimos ląstelės srities arba generuojama kitos ląstelės; 2) kinta ekstraląstelinė jonų koncentracija (dažnai [K + ] išeiti ); 3) atsiveria membraniniai jonų kanalai. Membranos potencialo pokyčiai gali turėti įtakos membranos baltymų konformacijai, todėl kanalai gali atsidaryti arba užsidaryti. Kaip aprašyta aukščiau, kai kurių membraninių siurblių veikimas priklauso nuo membranos potencialo. Nervų ląstelės yra specializuotos suvokti membranos potencialo pokyčius kaip informaciją, kuri turi būti apdorota ir perduodama (žr. 2 skyrių).
Ekstraląstelinės reguliuojančios medžiagos . Svarbiausias reguliavimo mechanizmas, susijęs su ekstraląstelinėmis medžiagomis, yra jų sąveika su specifiniais receptoriais plazmos membranoje arba ląstelės viduje. Šioms medžiagoms priskiriami sinapsiniai siųstuvai, perduodantys informaciją tarp nervinių ląstelių, vietiniai agentai ir medžiagos, kurios cirkuliuoja kraujyje ir pasiekia visas kūno ląsteles, pavyzdžiui, hormonai ir antigenai. Sinaptiniai siųstuvai yra mažos molekulės, išsiskiriančios iš nervų galūnėlių sinapsėje;
pasiekę kaimyninės, postsinapsinės ląstelės plazminę membraną, suveikia elektrinius signalus ar kitus reguliavimo mechanizmus. Šis klausimas išsamiai aptariamas skyriuje. 3.
Vietiniai cheminiai agentai dažnai išskiria specializuotos ląstelės. Jie laisvai difunduoja tarpląstelinėje erdvėje, tačiau jų veikimas apsiriboja nedidele ląstelių grupe dėl greito šių medžiagų sunaikinimo spontaniškai arba veikiant fermentams. Vienas iš tokių agentų išleidimo pavyzdžių yra išleidimas histaminas putliųjų ląstelių dėl pažeidimo ar imuninio atsako. Histaminas atpalaiduoja kraujagyslių lygiųjų raumenų ląsteles, padidina kraujagyslių endotelio pralaidumą ir stimuliuoja jutimo nervų galūnes, kurios sukelia niežulio pojūtį. Kitus vietinius cheminius veiksnius išskiria daugelis kitų ląstelių. Tipiški vietiniai agentai yra prostaglandinai, sudaro maždaug 20 riebalų rūgščių darinių grupę. Jie nuolat išsiskiria iš plačiai paplitusių ląstelių, tačiau veikia tik lokaliai, nes greitai sunaikinami membranų fosfolipazės. Įvairūs prostaglandinai pasižymi plačiu veikimo spektru: gali sukelti lygiųjų raumenų ląstelių susitraukimą, sukelti kraujo trombocitų (trombocitų) agregaciją arba slopinti geltonkūnio vystymąsi kiaušidėse.
Tarnauja kiti vietiniai agentai augimo faktoriai. Geriausiai žinomas yra simpatiniams neuronams skirtas nervų augimo faktorius (NGF), kuris būtinas šių neuronų augimui ir išlikimui vystymosi metu. in vivo arba ląstelių kultūroje. Matyt, šios neuronų klasės tikslinės ląstelės išskiria NGF ir taip užtikrina tinkamą inervaciją. Formuojant organus ląstelėms dažnai reikia „rasti kelią“ į tikslines ląsteles, kurios gali būti dideliais atstumais. Atitinkamai, turi būti daug specializuotų augimo faktorių, panašių į NGF.
Hormonai ir antigenai krauju pernešamos į visas ląsteles. Antigenai sukelia imuninį atsaką iš ląstelių, turinčių specifinių antikūnų. Tačiau antigenai, kaip taisyklė, yra svetimos medžiagos, kurios nesigamina reaguojančiame organizme (plačiau žr. 18 skyrių). Kai kurie hormonai, tokie kaip insulinas ar tiroksinas, veikia įvairius ląstelių tipus, o kiti, pavyzdžiui, lytiniai hormonai, veikia tik tam tikro tipo ląsteles. Hormonai yra arba peptidai, kurių veikimą sukelia prisijungimas prie ląstelės membranoje esančių receptorių, arba steroidai ir tiroksinas, kurie difunduoja per lipidų membraną ir jungiasi prie tarpląstelinių receptorių. Steroidiniai hormonai jungiasi prie branduolio chromatino, todėl transkripcija vyksta tam tikri genai. Gauti baltymai sukelia ląstelių funkcijų pokyčius, o tai yra specifinis hormonų poveikis. Su hormonų išsiskyrimu ir veikimu susiję klausimai išsamiai aptariami skyriuje. 17.
Tarpląstelinis ryšys, kuriame dalyvauja antrieji pasiuntiniai
Aukščiau aprašytos reguliavimo funkcijos apima poveikį ląstelės membranai. Informacija, kurią gauna ląstelės membrana, dažnai turi sukelti organelių reakciją ir yra perduodama joms įvairių medžiagų, vadinamų antraisiais pasiuntiniais (priešingai nei pirmieji pasiuntiniai, kurie į ląstelę patenka iš išorinių šaltinių). Antrųjų tarpininkų tyrimas sparčiai vystosi, todėl nėra garantijos, kad dabartinis problemos supratimo lygis bus pakankamai išsamus. Čia paliesime tris gerai ištirtus mediatorius: Ca 2+ , cAMP ir inozitolio trifosfatą.
Kalcis.Paprasčiausias tarpląstelinis tarpininkas yra Ca 2+ jonas. Jo laisvoji koncentracija ramybės būsenoje yra labai maža ir siekia 10_–8–10–7 mol/l. Jis gali patekti į ląstelę tam tikrais membranos kanalais, kai jie yra atviroje būsenoje, pavyzdžiui, pasikeitus membranos potencialui (žr. 2 skyrių). Dėl to didėjanti Ca 2+ koncentracija ląstelėje sukelia svarbias reakcijas, tokias kaip miofibrilių, kurios yra raumenų susitraukimo pagrindas (žr. 4 skyrių), arba pūslelių, turinčių neuromediatorių, išsiskyrimą iš nervų galūnių (žr. 3 skyrių). . Abiem reakcijoms reikalinga maždaug 10–5 mol/l Ca 2+ koncentracija. Reguliuojamąjį poveikį turintis Ca 2+ gali išsiskirti ir iš tarpląstelinių atsargų, tokių kaip endoplazminis tinklas. Ca 2+ išleidimui iš depo būtinas kitų tarpininkų dalyvavimas (žr., pvz., 1.16 pav.).
Ciklinis adenozino monofosfatas, cAMP. Neseniai buvo įrodyta, kad ciklinis adenozino monofosfatas (cAMP), pagrindinio organizmo energijos šaltinio ATP darinys, yra svarbus antrasis pasiuntinys. Sudėtinga reakcijų grandinė, parodyta Fig. 1.15, prasideda nuo receptorių R s ant išorinio plazminės membranos paviršiaus, kuris gali būti specifinė įvairių mediatorių ir hormonų surišimo vieta. Prisijungus prie konkrečios „stimuliuojančios“ molekulės R s keičia savo formą; šie pokyčiai veikia baltymus G s ant vidinio membranos paviršiaus taip, kad pastarąją būtų galima suaktyvinti viduląsteliniu guanozino trifosfatu (GTP). Aktyvuotas baltymas G s , savo ruožtu, stimuliuoja vidiniame membranos paviršiuje esantį fermentą adenilato ciklazę (AC), kuri katalizuoja cAMP susidarymą iš ATP. Vandenyje tirpus cAMP yra tarpininkas, perduodantis poveikį

Ryžiai. 1.15.Reakcijų grandinė, apimanti tarpląstelinį pasiuntinį cAMP (ciklinį adenozino monofosfatą). Jaudinantys arba slopinantys išoriniai signalai aktyvuoja membranos receptorius R s arba Ri . Šie receptoriai reguliuoja jungimosi procesą G – baltymai, turintys tarpląstelinį GTP (guanozino trifosfatą), taip stimuliuojantys arba slopinantys intracelulinę adenilato ciklazę (AC). Stiprinantis fermentas AC paverčia adenozino trifosfatą (ATP) į cAMP, kurį vėliau fosfodiesterazė suskaido į AMP. Laisvas cAMP difunduoja į ląstelę ir aktyvuoja adenilatkinazę (A-kinazę), išskirdamas jos katalizinį subvienetą C, kuris katalizuoja viduląstelinių baltymų fosforilinimą, t.y. formuoja galutinį ekstraląstelinio dirgiklio poveikį. Diagramoje taip pat rodomi farmakologiniai vaistai ir toksinai, kurie sukelia (+) arba slopina (-) tam tikras reakcijas (modifikuotas)
ekstraląstelinių receptorių stimuliavimas R s į vidines ląstelės struktūras.
Lygiagrečiai su stimuliuojančia reakcijų grandine, apimančia R s galimas slopinančių mediatorių ir hormonų prisijungimas prie atitinkamų receptorių R i kuris vėlgi per GTP aktyvuotą baltymą G , slopina AC, taigi ir cAMP gamybą. Pasklidęs į ląstelę, cAMP reaguoja su adenilatkinaze (A-kinazė); tai išleidžia C subvienetą, kuris katalizuoja baltymo P fosforilinimą. Dėl šio fosforilinimo baltymai paverčiami aktyvia forma ir dabar jie gali turėti specifinį reguliuojantį poveikį (pavyzdžiui, sukelti glikogeno skaidymą). Ši sudėtinga reguliavimo sistema yra labai efektyvi, nes galutinis rezultatas yra daugelio baltymų fosforilinimas, ty reguliavimo signalas keliauja per grandinę su dideliu stiprėjimu. Išoriniai mediatoriai, kurie jungiasi prie receptorių R s ir R i kiekvienam iš jų būdingas, itin įvairus. Adrenalinas, jungiantis su R s arba R i dalyvauja reguliuojant lipidų ir glikogeno apykaitą, taip pat stiprinant širdies raumens susitraukimą ir kitose reakcijose (žr. 19 skyrių). Skydliaukę stimuliuojantis hormonas, aktyvuojantis R s , skatina skydliaukės hormono tiroksino išsiskyrimą, o prostaglandinas I slopina kraujo trombocitų agregaciją. Slopinamasis poveikis, įskaitant adrenaliną, tarpininkauja R i yra išreikšti lipolizės sulėtėjimu. Taigi, cAMP sistema yra daugiafunkcinė tarpląstelinė reguliavimo sistema, kurias gali tiksliai kontroliuoti tarpląstelinės stimuliuojančios ir slopinančios signalinės medžiagos.
Inozitolio fosfatas "IF" h “ Antrojo pasiuntinio – inozitolio fosfato – tarpląstelinė sistema buvo atrasta visai neseniai (1.16 pav.). Šiuo atveju nėra slopinimo kelio, tačiau yra panašumas į cAMP sistemą, kurioje R receptoriaus stimuliacijos poveikis perkeliamas į GTP aktyvuotą G baltymą, esantį vidiniame membranos paviršiuje. Kitame etape įprastas membraninis lipidas fosfatidilinozitolis (PI), prieš tai gavęs dvi papildomas fosfatų grupes, paverčiamas PI-difosfatu (PIP 2), kurį aktyvuota fosfodiesterazė (PDE) suskaido į inozitolio trifosfatas(IFz) ir lipidų diacilglicerolis(DAG). Inozitolio trifosfatas yra vandenyje tirpus antrasis pasiuntinys, kuris difunduoja į citozolį. Jis pirmiausia veikia išskirdamas Ca 2+ iš endoplazminio tinklo. Ca 2+ savo ruožtu veikia kaip pasiuntinys, kaip aprašyta aukščiau; pavyzdžiui, aktyvuoja nuo Ca 2+ priklausomą fosfokinazę, kuri fosforilina fermentus. DAG lipidinis subvienetas (1.16 pav.) taip pat neša signalą, plazmos membranos lipidinėje fazėje difunduodamas į C-kinazę, esančią jos vidiniame paviršiuje, kuri aktyvuojama dalyvaujant fosfatidilserinui kaip kofaktoriui. Tada C-kinazė sukelia baltymų fosforilinimą, paverčiant juos aktyvia forma.
Intraląstelinė antrojo pasiuntinio sistema IF 3 taip pat gali būti kontroliuojama įvairių išorinių mediatorių ir hormonų, įskaitant acetilcholiną, serotoniną, vazopresiną ir skydliaukę stimuliuojantį hormoną; kaip ir cAMP sistema, jai būdingas įvairus tarpląstelinis poveikis. Gali būti, kad šią sistemą taip pat suaktyvina šviesa akies regos receptoriuose ir atlieka pagrindinį vaidmenį fototransdukcijoje (žr. 11 skyrių). Pirmą kartą individualioje organizmo raidoje IG sistemos receptorius aktyvuoja spermatozoidai, dėl to IG dalyvauja reguliacinėse reakcijose, lydinčiose kiaušialąstės apvaisinimą.
cAMP ir IFz-DAG sistemos yra labai veiksmingos biologiniai stiprikliai. Jie

Ryžiai. 1.16.Reakcijų grandinė, kurioje dalyvauja intracelulinis tarpininkas IPG (inozitolio trifosfatas). Kaip ir cAMP sistemoje, tarpląstelinis signalas yra perduodamas per baltymąG, kuri šiuo atveju aktyvuoja fosfodiesterazę (PDE). Šis fermentas skaido fosfatidilinozino difosfatą (PIP 2 ) plazmos membranoje prieš IF h ir diacilglicerolis (DAG); JEI h difunduoja į citoplazmą. Čia tai sukelia Ca išsiskyrimą 2+ iš endoplazminio tinklo; Ca koncentracijos padidėjimas 2+ citoplazmoje ([Ca 2+ ]i ) aktyvina proteinkinazę, kuri fosforilina, todėl aktyvina fermentus. Kitas produktas, DAG, lieka membranoje ir aktyvuoja proteinkinazę C (fosfatidilserino kofaktorius, PS). Baltymų kinazė C taip pat fosforilina fermentus, kurie tarpininkauja specifiniams veiksmams, susijusiems su išorinių receptorių stimuliacija R . Reakcijų grandinės, susijusios su IF, atšakos h ir DAG gali būti aktyvuojami nepriklausomai atitinkamai jonomicinu ir forbolio esteriu (modifikuotas)
paverčia reakciją tarp siųstuvo ir išorinės membranos receptoriaus į įvairių tarpląstelinių baltymų fosforilinimą, o tai gali paveikti įvairias ląstelių funkcijas. Vienas iš reikšmingų problemos aspektų yra tai, kad, kiek šiandien žinoma, yra tik šios dvi glaudžiai susijusios tokio tipo reguliavimo sistemos, kurias naudoja daugybė išorinių tarpininkų, reguliuojančių įvairius tarpląstelinius procesus. Tuo pačiu metu šios reguliavimo sistemos, įskaitant Ca 2+, glaudžiai sąveikauja viena su kita, o tai leidžia joms tiksliai reguliuoti ląstelių funkcijas.
Mokymai ir vadovai
1. Alberts IN., Bray D., Lewis J., Raff M., Roberts KAM., Watson J.D.Ląstelės molekulinė biologija, Niujorkas ir Londonas, Garland Publishing Inc., 1983 m.
2. Czihak G., Longer H., Ziegler H.(red.). Biologija. Berlynas, Heidelbergas, Niujorkas, Springeris, 1983 m.
3. Hille IN. Jaudinamųjų membranų joniniai kanalai. Sanderlandas, Masažas, Sinauer Assoc., 1984 m.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(red.). Biofizika. Berlynas, Heidelbergas, Niujorkas, Springeris, 1984 m.
5. Jungermannas KAM., Mahleris H.Biochemija. Berlynas, Heidelbergas, Niujorkas, Springeris, 1980 m.
6. Kandel E. R., Schwartz – J. H.,(red.). Neuronologijos principai, Niujorkas, Amsterdamas, Oksfordas, Elsevier, 1985 m.
7. Šibeleris T. H., Schmidtas W.Menscheno anatomija. Berlynas, Heidelbergas, Niujorkas, Tokijas, Springeris, 1983 m.
Originalūs straipsniai ir apžvalgos
8. Berridge M.J. Molekulinis bendravimo ląstelėje pagrindas, Sci. Amer, 253, 124-134 (1985).
9. Berridge M. J., Irvine R. F. Inozitolio trifosfatas, naujas antrasis korinio signalo perdavimo pasiuntinys. Nature, 312, 315, 321 (1984).
10. Bretscher M.S. Ląstelės membranos molekulės, Sci. Amer., 253, 124–134 (1985).
11. Dautas J. Gyva ląstelė kaip energiją perduodanti mašina. Minimalus miokardo metabolizmo modelis, Biochem. et Biophys. Acta, 895, 41–62 (1987).
12. Hodžkinas A.L., Katz IN. Natrio jonų poveikis milžiniško kalmarų aksono elektriniam aktyvumui. J. Physiol. (Lond.), 108, 37–77 (1949).
13. Hodžkinas A.L., Keynesas R.D. Aktyvus katijonų pernešimas milžiniškuose aksonuose iš Sepija ir Loligo, J. Physiol. (Lond.), 128, 28–42 (1955).
14. Ilgesnis P. Jonų kanalai su konformaciniais subbūsenais, Biophys. J., 47, 581–590 (1985).
15. Ochs S., vertas P.M. Egzoplazminis transportas normaliose ir patologinėse sistemose. In: Physiology and Pathology of Axons, S.G. Waxmann, Red. Niujorkas, Raven Press, 1978 m.



